К. Э. Фабри
Основы зоопсихологии

3-е издание
Рекомендовано Министерством общего и профессионального образования Российской Федерации в качестве учебника для студентов высших учебных заведений, обучающихся по специальностям «Психология», «Биология», «Зоология» и «Физиология»
Москва
Российское психологическое общество
1999
ББК88.2
Ф12
Рекомендовано к изданию Ученым Советом факультета психологии Московского государственного университете имени М. В. Ломоносова
Рецензенты:
Л. И. Анцыферова, доктор психологических наук, кафедра зоологии позвоночных животных и общей экологии биологического факультета Московского государственного университета имени М. В. Ломоносова
Фабри К.Э.
Основы зоопсихологии: Учебник для студентов высших учебных заведений,
Ф 12 обучающихся по специальностям «Психология», «Биология», «Зоология» и «Физиология». – 3-е изд. – М.: Российское психологическое общество, 1999. –464 с.
ISBN 5-89573-051-5
Учебник (1-е изд. – 1976 г., 2-е изд. – 1993 г.), написанный видным зоопсихологом К. Э. Фабри, посвящен возникновению, развитию и функционированию психики у животных. Освещаются проблемы общей психологии: отражательная природа психики, взаимосвязь психики и поведения, соотношение врожденного и приобретенного, закономерности развития психики в филогенезе, условия и предпосылки возникновения и развития психики человека. Дается широкое обобщение и анализ современных достижений этологических и зоопсихологических исследований. Приводятся результаты многочисленных эмпирических исследований.
Для студентов высших учебных заведений, обучающихся по специальностям «Психология», «Биология», «Зоология» и «Физиология», а также для всех, интересующихся поведением и психикой животных.
ББК88.2
ISBN 5-89573-051-5 © Фабри К.Э.,1973, 1993,1999.
© Российское психологическое
общество, 1999.
Предисловие к 1-му изданию
Зоопсихология является неотъемлемой и важной частью психологических наук. Интерес к ней заметно вырос в последние годы, особенно в связи с тем, что в этой области знаний сейчас наблюдается большой подъем исследовательской работы и творческой мысли. Наряду с большим теоретическим значением зоопсихологических исследований все более расширяется и сфера их практического использования. Плодотворная работа исследователей многих стран по изучению поведения животных, охватывающая все зоологические таксоны – от амебы до шимпанзе, – привела к весьма внушительным результатам: в настоящее время накоплено такое обилие ценнейших научных фактов, что не только удалось разъяснить ряд существенных вопросов зоопсихологии, но и пришлось внести серьезные поправки в, казалось бы, вполне устоявшиеся положения и концепции. Разумеется, и поныне остается еще очень много неизученного и непонятного в процессах поведения – этих сложнейших природных процессах. Все же мы можем сейчас лучше представить биологические предпосылки и предысторию зарождения человеческого сознания.
Зоопсихология входит как обязательный курс в программу обучения на факультетах психологии университетов, но учебника зоопсихологии до сих пор не существовало ни в Советском Союзе, ни за рубежом. В последние годы, правда, появились монографии и обзорные работы по поведению животных, но эти публикации носят лишь этологический характер, и в них совсем не затрагиваются вопросы, связанные с психическим аспектом поведения животных, т.е. те вопросы, изучение которых как раз важно для студента-психолога.
Данная книга написана на основе многолетнего опыта автора по чтению курсов лекций по зоопсихологии и этологии в Московском университете. Чисто этологический материал дается в ней лишь в той мере, в какой это необходимо для уяснения зоопсихологических вопросов. При этом делается упор на освещение развития психики животных как в онто-, так и в филогенезе. Ведь зоопсихология – это наука о становлении и развитии психики на дочеловеческом уровне, и познать психику животных вне процесса их развития принципиально невозможно. В заключительной главе очень кратко излагаются и некоторые моменты, касающиеся непосредственной предыстории антропогенеза. Конкретный фактический материал, использованный в тексте, относится преимущественно к исследованиям последних 5–10 лет, но приводятся и важные исследования давних лет, преимущественно советских ученых.
Первая попытка создания учебника зоопсихологии не может оказаться без недостатков, и автор заранее выражает свою благодарность за критические замечания.
Предисловие ко 2-му изданию*
* 2-е издание учебника подготовила полностью к публикации ученица К.Э.Фабри – кандидат психологических наук Н.Н.Мешкова.
«Основы зоопсихологии» К. Э. Фабри, впервые изданные в 1976 г., сохраняют свою актуальность и научную ценность и сегодня.
Читатель книги может узнать, есть ли у животных психика, каковы ее особенности, как ее изучать, что такое поведение, как оно связано с психикой, где используются зоопсихологические знания, что такое зоопсихология как наука и как она связана с общей психологией и биологией.
Книга дает также важный материал для понимания общей природы психического. Только зная психику животных, мы можем в человеческой психике различать общее для человека и животных и специфическое только для человека или только для животных, избежать ошибок антропоморфизма, биологизаторства, физиологического редукционизма, подмены родовых характеристик психического теми, которые свойственны только человеку. В этом бесспорный вклад зоопсихологии в общую психологическую культуру.
«Основы» имеют четкую теоретическую ориентацию на диалектический материализм, на диалектико-материалистическую психологию. Вслед за ведущими теоретиками общей психологии К. Э. Фабри считает категорию «психическое отражение» центральной и анализирует его особенности, функционирование и развитие.
Сегодня в отечественной психологии преодолевается односторонняя ориентация на философию диалектического материализма. Думаю, что это затронет и зоопсихологию. Более полного представления требуют знания о мотивационно-эмоциональной сфере в психике животных.
«Основы» отражают определенный этап в развитии зоопсихологии и дают многое для ее последующего обогащения. Последние работы К.Э.Фабри (см. Приложение), которые дополняют текст первого издания учебника, свидетельствуют о том, что сам он постоянно думал о дальнейшем развитии науки о психике животных.
O.K. Тихомиров, профессор Февраль 1992 г.
К.Э.Фабри и зоопсихология в Московском государственном университете имени М. В. Ломоносова
Автор лежащего перед вами учебника – профессор факультета психологии МГУ К.Э.Фабри (1923–1990), один из ведущих зоопсихологов нашей страны. К.Э.Фабри родился в Вене, в 1932 г. он вместе с родителями переехал в Советский Союз, где его отцу, видному деятелю Компартии Австрии, было предоставлено политическое убежище. Перед самой войной К.Э.Фабри поступил на биологический факультет МГУ, но учеба была прервана войной, в течение которой он работал сначала в госпиталях, а затем военным переводчиком. Вернувшись после войны на биофак МГУ, Курт Эрнестович стал одновременно специализироваться по зоологии позвоночных и антропологии, а также посещал занятия на отделении психологии философского факультета МГУ. Его дипломная работа, посвященная сложным формам поведения обезьян, получила особую, даваемую крайне редко оценку – «отлично с отличием» и право на публикацию.
Научным руководителем К.Э.Фабри и в студенческие годы и в годы его первых самостоятельных исследований была Надежда Николаевна Ладыгина-Котс – ведущий отечественный зоопсихолог. Именно у нее он учился тому, как нужно планировать и проводить зоопсихологический эксперимент, вести наблюдения за поведением животных. К сожалению, зоопсихология в 40–60 гг. в Советском Союзе была объявлена, наряду с генетикой и кибернетикой, «лженаукой», и в целом разделила с ними их печальный путь. Однако, К.Э.Фабри всегда оставался верным выбранному пути, и, хотя жизнь и заставила его после окончания МГУ работать то в Библиотеке иностранной литературу, то в Радиокомитете, он продолжал заниматься «неофициальной» научной работой – изучением манипуляционных особенностей разных видов млекопитающих и, в особенности, обезьян. Н. Н. Ладыгина-Котс, сама находившаяся в это время в «научном подполье», оставалась для него наставником и другом.
Только в 1964 г., через 18 лет после окончания МГУ, К.Э.Фабри получил возможность официально заняться изучением поведения и психики животных. Для этого ему пришлось уехать в г. Пущино-на-Оке, где в то время был организован Академгородок. К.Э.Фабри поступил на работу в Институт биофизики, здесь им были выполнены оригинальные исследования импринтинга у птиц, существенно дополнившие классические представления о природе этого феномена.
В 1966 г. А. Н. Леонтьев пригласил К.Э.Фабри для чтения лекций по зоопсихологии и этологии на только что организованном факультете психологии МГУ. Много раньше этот курс в Университете читала Н. Н. Ладыгина-Котс, с началом «оттепели» это почетное право получил ее ближайший ученик. С 1971 г. кафедра общей психологии становится местом его постоянной работы, сначала в качестве старшего преподавателя, затем доцента, а с 1983 г. – профессора. За эти годы сотни студентов прослушали его курс «Основы зоопсихологии», а также спецкурс по этологии, многие выполнили под его руководством курсовые и дипломные работы, защитили кандидатские диссертации. К.Э.Фабри – автор более 200 научных публикаций. Основная тематика его работ, значительная часть которых выполнена в стенах факультета психологии, раскрывает различные аспекты онтогенеза поведения и психики животных, эволюции психики, психической деятельности приматов, этологические и биопсихологические предпосылки антропогенеза. В 1977 г. К.Э.Фабри за написание учебника «Основы зоопсихологии» была присуждена степень доктора психологических наук. С того времени эта книга выдержала 3 переиздания (первые два – в 1976 и 1993 гг.), оставаясь и до сих пор основным и практически единственным учебником по зоопсихологии для студентов высших учебных заведений.
Следует упомянуть еще одну заслугу К.Э.Фабри перед отечественными читателями. Именно благодаря его усилиям труды К. Лоренца и Н. Тинбергена, с которыми, кстати, он вел научную переписку, стали широко доступны в нашей стране. Именно К.Э.Фабри сумел доказать необходимость перевода книг выдающихся этологов, в последствие ставших лауреатами Нобелевской премии, был научным редактором и автором предисловий первых советских изданий их книг.
В 1977 г. при поддержке А. Н. Леонтьева на факультете психологии МГУ была организована небольшая, состоящая всего из трех сотрудников, группа зоопсихологов, которая и стала началом будущей лаборатории зоопсихологии. За прошедший с того времени период из стен лаборатории вышли специалисты, нашедшие приложение свои знаниям в области психологии и поведения животных в научных учреждениях АН СССР и России, в заповедниках, в институтах и научных центрах Академии педагогических наук СССР и России. Было защищено несколько диссертаций, посвященных ориентировочно-исследовательской деятельности животных, исследованию мотивации игр животных, сравнительному анализу манипуляционной активности разных видов млекопитающих, онтогенезу интеллекта антропоидов.
В настоящее время лаборатория зоопсихологии при кафедре общей психологии продолжает работать по-прежнему небольшим составом, но тематика исследований в соответствии с требованиями сегодняшнего дня несколько поменялась. Наряду с проблематикой, относящейся к области фундаментальных исследований – изучением психологических закономерностей и механизмов, лежащих в основе антропогенной эволюции высших позвоночных животных (итогом работы по этой тематике стала монография Н. Н. Мешковой и Е. Ю. Федорович «Ориентировочно-исследовательская деятельность, подражание и игра как психологические механизмы адаптации высших позвоночных к урбанизированной среде»- М.: Аргус, 1996), появились и темы прикладного характера. Следует отметить, что начало прикладным исследованиям было положено еще раньше, когда лабораторией руководил К.Э.Фабри. Тогда, в 1985–1989 гг., был проведен цикл исследований поведения рыб в зоне промышленного лова (по заказу самих специалистов в области промышленного рыболовства), ставших в определенной степени основой для перестройки самой стратегии создания орудий лова. Но, пожалуй, важнее, что эта работа способствовала изменению отношения к самой рыбе – объекту лова – как к животному, обладающему достаточно высоким уровнем развития перцептивной психики, способному тонко адаптироваться к условиям промысла. Как итог этой работы было выпущено пособие по их тиопсихологии, написанное К.Э.Фабри по заказу Калининградского технического института рыбной промышленности и хозяйства, а также разработан курс лекций для студентов-промысловиков.
Прикладные темы, разрабатываемые в настоящее время в лаборатории зоопсихологии, посвящены психологическим аспектам взаимоотношений горожанина с домашними животными, а также исследованию становления отношений «ребенок–животное».
Лаборатория зоопсихологии продолжает и активную педагогическую деятельность. Сотрудники лаборатории читают курсы лекций для студентов и слушателей факультета психологии МГУ, ведут практикум по теме «Психологическое наблюдение за животными». В 1997 г. была подготовлена и издана хрестоматия по зоопсихологии и сравнительной психологии.
Н.Н.Мешкова, кандидат психологических наук, старший научный сотрудник, Е. Ю. Федорович, младший научный сотрудник (Лаборатория зоопсихологии кафедры общей психологии факультета психологии Московского государственного университета имени М. В. Ломоносова)
Введение
Предмет и задачи зоопсихологии
Среди великих загадок природы, к познанию которых с древнейших времен стремился человеческий ум, психика животных занимает одно из первых мест. «Душевная жизнь», психические качества и поведение животных входили как существенные составные части в фольклор, религиозные представления и мировоззрение на всех этапах развития человечества. Много внимания этим вопросам, особенно отношению психики животных к психике человека, уделяли античные мыслители при построении своих философских концепций.
А.И.Герцен писал: «Психология животных несравненно менее обращала на себя внимание ученых-естествоиспытателей, нежели их форма. Животная психология должна завершить, увенчать сравнительную анатомию и физиологию; она должна представить дочеловеческую феноменологию развертывающегося сознания; ее конец – при начале психологии человека, в которую она вливается, как венозная кровь в легкие, для того, чтобы одухотвориться и сделаться алой кровью, текущею в артериях истории».*
* Герцен А.И. Избранные философские сочинения. М., 1940. С. 222–223.
В наше время изучением психики животных занимается специальная наука – зоопсихология. Но прежде чем приступить к рассмотрению предмета и задач зоопсихологии, необходимо уточнить, что мы понимаем под психикой, поведением и психической деятельностью у животных.
Согласно ленинской теории отражения психика является по своему содержанию высшей формой отражения объективной реальности. Появившись лишь на определенном этапе развития органического мира, психика присуща только высокоорганизованным живым существам. Она выражается в их способности отражать своим состоянием окружающий мир. Началом этого этапа в эволюции органического мира следует считать появление животной формы жизни, ибо именно специфические условия жизнедеятельности животных породили необходимость качественно нового, активного отражения объективной действительности, способного регулировать усложнившиеся отношения организма со средой.
Таким образом, психика является формой отражения, позволяющей животному организму адекватно ориентировать свою активность по отношению к компонентам среды. При этом, служа активному отражению объективной реальности, материи, психика сама есть свойство высокоразвитой органической материи. Этой материей является нервная ткань животных (или ее аналоги). У подавляющего большинства животных имеется головной мозг – центральный орган нервно-психической деятельности.
Психика животных неотделима от их поведения, под которым мы понимаем всю совокупность проявлений внешней, преимущественно двигательной активности животного, направленную на установление жизненно необходимых связей организма со средой. Психическое отражение осуществляется на основе этой активности в ходе воздействий животного на окружающий мир. При этом отражаются не только сами компоненты окружающей среды, но и собственное поведение животного, а также произведенные им в результате этих воздействий изменения в среде. Притом у высших животных (у высших позвоночных), которым свойственны подлинные познавательные способности, наиболее полноценное и глубокое отражение предметов окружающего мира совершается именно в ходе их изменения под воздействием животного.
Таким образом, справедливо считать психику функцией животного организма, состоящей в отражении предметов и явлений окружающего мира в ходе и результате направленной на этот мир активности, т.е. поведения. Внешняя активность и ее отражение, поведение и психика составляют неразрывное органическое единство и могут лишь условно расчленяться для научного анализа. Как показал еще И.М.Сеченов, психика зарождается и умирает с движением, поведением.
Итак, первопричина психического отражения – поведение, посредством которого осуществляется взаимодействие с окружающей средой, без поведения нет психики. Но справедливо и обратное, ибо, являясь производной поведения, психика вторично сама корректирует и направляет внешнюю активность организма. В этом и состоит приспособительная роль психики: адекватно отражая окружающий мир, животное приобретает возможность ориентироваться в нем и в результате адекватно строить свои отношения с биологически значимыми компонентами среды.
Сущность диалектического единства поведения и психики лучше всего выражает понятие «психическая деятельность». Под психической деятельностью животных мы понимаем весь комплекс проявлений поведения и психики, единый процесс психического отражения как продукт внешней активности животного. Такое понимание психической деятельности, неразрывного единства психики и поведения животных, открывает перед зоопсихологией путь к истинному познанию их психических процессов и к плодотворному изучению путей и закономерностей эволюции психики. Поэтому, учитывая примат поведения в психическом отражении, мы будем при обсуждении отдельных аспектов психической деятельности животных исходить прежде всего из анализа их двигательной активности в конкретных условиях их жизни.
Определив объект зоопсихологии – психическую деятельность животных, мы можем теперь сформулировать предмет зоопсихологии как науки о проявлениях, закономерностях и эволюции психического отражения на уровне животного, о происхождении и развитии в онто- и филогенезе психических процессов у животных и о предпосылках и предыстории человеческого сознания. Зоопсихолог изучает эволюцию психики начиная с ее зачаточных форм до высших ее проявлений, составлявших основу зарождения человеческой психики.
Таким образом, компетенция зоопсихолога находится в пределах двух граней – нижней и верхней, представляющих собой одновременно главные вехи эволюции психики вообще. Нижняя грань знаменует начало психического отражения, исходную ступень его развития, верхняя – смену животной психики человеческой. Нижняя грань означает проблему качественного различия отражения у растений и животных, верхняя – у животных и людей. В первом случае приходится решать вопросы происхождения психики из более элементарной формы отражения, во втором – зарождения человеческой психики из элементарной по отношению к ней психики животных.
Материалистическое решение проблемы зарождения психики возможно лишь на основе признания возникновения психики из раздражимости, свойственной всей живой материи, в которой уже, как писал выдающийся советский психолог Л.С.Выготский, «заложены зачатки того, из чего впоследствии путем длительной эволюции должна развиваться человеческая психика».* Перефразируя изречение В.Гёте о том, что «все растения произошли из одного», Выготский подчеркивал, что «все формы психических явлений произошли из раздражимости».**
* Выготский Л.С. Развитие высших психических функций. М., 1960. С. 405.
** Там же.
Основательную разработку проблема зарождения и развития психики получила в исследованиях известного советского психолога A. Н. Леонтьева. Леонтьев исходит из того, что живая материя на допсихическом уровне своего развития характеризуется в функциональном отношении «простой раздражимостью», т.е. способностью отвечать избирательно и специфическим образом на жизненно значимые воздействия среды в соответствии с потребностями обмена веществ. Биологическая эволюция сопряжена с усложнением обмена веществ. Соответственно взаимоотношения организма со средой развиваются в сторону все большего разнообразия видов раздражимости, так как организмы ассимилируют из внешней среды все большее число веществ и форм энергии.
Однако, как указывает Леонтьев, развитие раздражимости не сводится лишь к такому количественному усложнению. В результате того, что в ходе эволюции меняется также общий тип взаимодействия организмов со средой, возникает качественно новая форма раздражимости – чувствительность. Чувствительность же – это способность к ощущению, т.е. элементарное психическое явление. Другими словами, психика начинается там, где появляется качественно своеобразная высшая форма раздражимости – собственно чувствительность.
Важно подчеркнуть, что появление чувствительности знаменуется, как пишет Леонтьев, тем, что «организмы становятся раздражимыми и по отношению к таким воздействиям, которые сами по себе не в состоянии определить ни положительно, ни отрицательно их ассимилятивную деятельность, обмен веществ с внешней средой».*
* Леонтьев А.Н. Проблемы развития психики. 3-е изд. М., 1972. С. 43.
Леонтьев поясняет это положение следующим примером. Лягушка раздражима по отношению к такому воздействию, как шорох, поскольку он вызывает у нее определенную (ориентировочную) реакцию. Но при этом «энергия звука шороха, воздействующая на организм лягушки, ни на одной из ступеней своего преобразования в организме не ассимилируется им и вообще прямо не участвует в его ассимилятивной деятельности. Иначе говоря, само по себе данное воздействие не может служить поддержанию жизни организма и, наоборот, оно вызывает лишь диссимиляцию вещества организма».* Биологическая же роль подобных воздействий и их отражения в виде ощущений заключается в том, что эти сами по себе непосредственно жизненно незначимые воздействия становятся для животного сигналами о возможности появления и поглощения необходимого для поддержания жизни вещества или энергии (в данном примере – насекомого и т.п.).
* Там же. С. 44.
Таким образом, заключает Леонтьев, новая форма раздражимости, отражающая подобные воздействия, «опосредствует деятельность организма, направленную на поддержание жизни».* Наступает как бы раздвоение прежде единого процесса взаимодействия организма с внешней средой: «С одной стороны, выделяются процессы, с которыми непосредственно связаны поддержание и сохранение жизни. Эти процессы составляют первую, исходную форму жизнедеятельности организмов. В ее основе лежат явления первичной раздражимости организмов.
* Там же.
С другой стороны, выделяются процессы, прямо не несущие функции поддержания жизни и лишь опосредствующие связи организма с теми свойствами среды, от которых зависит его существование. Они составляют особую форму жизнедеятельности, которая и лежит в основе чувствительности организмов, психического отражения ими свойств внешней среды».*
* Там же. С. 49–50.
В этой концепции генезиса и сущности чувствительности как первичной формы психического отражения, разработанной Леонтьевым, особенно существенным является то, что психическое качество выводится непосредственно из жизнедеятельности организма. Речь идет о раздвоении деятельности организма, его активности, причем психика появляется здесь как новое свойство внешней активности. При этом, естественно, сама внешняя активность претерпевает глубокие качественные преобразования и превращается в поведение.
Можно, очевидно, считать, что появление вместе с животной формой жизни поведения знаменуется переходом от неопосредованной (допсихической) внешней активности к активности, опосредованной отражением предметной действительности (т.е. психическим отражением).
Таким образом, поле деятельности зоопсихолога начинается у стыка допсихического и психического отражения, на уровне первых проявлений чувствительности, способности к ощущению у низших представителей животного мира. Отсюда берет свое начало эволюция психики. Поднимаясь как бы террасами, образуемыми качественными эволюционными преобразованиями животной психики, поле деятельности зоопсихолога простирается отсюда до рубежа человеческого сознания.
Определяя «верхнюю грань» зоопсихологических исследований, важно подчеркнуть, что верный путь к изучению этой проблемы открывается лишь при учете как общих с животными эволюционных корней психической деятельности животных и человека, так и качественных перестроек, поднявших эту деятельность на совершенно новый, небывалый до того уровень развития.
По сравнению с психикой животных психика человека представляет собой качественно иную категорию, хотя генетически и связанную с психикой животных. В поведении человека продолжают играть определенную роль биологические, общие с животными факторы, но сущность человеческого поведения определяется глубокими качественными различиями между человеком и животным. Эти различия обусловлены общественно-трудовой деятельностью и членораздельной речью, которые всецело отсутствуют у животных.
Выдающийся советский ученый, основоположник отечественной зоопсихологии В.А. Вагнер еще полвека тому назад всесторонне проанализировал взаимоотношения биологических и социальных факторов в жизни человека. Вагнер не отрицал наличия общих элементов в психике человека и животных, но подчеркивал как важнейшую задачу необходимость выявления и учета качественных различий между человеком и животными. Характерный для него истинно эволюционный подход к проблемам психической деятельности животных не имел ничего общего с плоским эволюционизмом, стирающим всякую грань между животным и человеком и постулирующим всю эволюцию психики от простейших до человека как цепь лишь количественных совершенствований и преобразований. Этот «монизм снизу» (по терминологии Вагнера), представленный разными вариантами в теории Ж.Леба о тропизмах, в положениях бихевиористов, рефлексологов и других, подвергся Вагнером острой критике.
Но, выступая против упрощенчества в толковании поведения животных, против низведения сложного к простому, низведения высших форм психических функций, в том числе и психики человека, к низшим, Вагнер одновременно вел не менее решительную борьбу с, казалось бы, противоположным течением – «монизмом сверху», с антропоморфизмом в зоопсихологии.
Антропоморфизм выражается в суждении о психических явлениях у животных по аналогии с таковыми у человека, в представлении о наличии у животных таких психических свойств и способностей, которые на самом деле присущи только человеку. Антропоморфическое толкование поведения животных с точки зрения человеческих мотивов и поступков, стирание грани между человеком и животным ведут к отрицанию качественных особенностей человеческой психики.
Сопоставляя «монизм сверху» и «монизм снизу», нетрудно заметить, что они, по существу, сливаются на базе общего отрицания качественных различий между отдельными этапами филогенеза и особенно между животным и человеком.
Характеризуя общее и различное в поведении человека и животных, Выготский указал на «то обстоятельство, что развитие высших психических функций происходит без изменения биологического типа человека, в то время, как изменение биологического типа является основой эволюционного типа развития».* Главное отличие психической деятельности человека от таковой у животных состоит в том, что благодаря общественно-трудовой практике и связанной с ней членораздельной речи человек способен к отражению объективной реальности в двух планах – чувственном и понятийном, т.е. наряду с непосредственно-чувственным отражением у него существует еще план абстрактно-логического мышления, осуществляемого с помощью понятий. Именно этим определяется сущность сознания.
* Выготский Л.С. Развитие высших психических функции. М., 1960. С. 40–41.
Что же касается животных, то даже у наиболее высокоразвитых их представителей психическое отражение окружающего мира совершается лишь в одном плане, а именно чувственном; второй, абстрактно-логический план у них отсутствует.
Однако без логического мышления (с помощью абстрактных понятий) невозможно постигнуть сущность вещей и процессов, невозможно познать коренные причинно-следственные связи, подлинные, существенные закономерности. Вследствие этого познание животными компонентов среды и отношений между ними ограничивается лишь непосредственно воспринимаемыми (в настоящем или прошлом), причем в наибольшей мере это познание осуществляется в ходе направленной на эти компоненты двигательной активности. Дистантное восприятие обычно дает животному лишь поверхностную, сигнальную информацию. Отражение же собственных действий, собственного поведения, направленного на объекты окружающего мира, дает животному наиболее полноценную информацию об этом мире. (В ограниченной степени познавательная деятельность встречается также в форме подражания, когда имеет место научение путем созерцания действий других животных.)
Человек же способен проникнуть в сущность вещей и явлений, познать закономерности их происхождения и развития, по определению В. И. Ленина, идя от живого созерцания к абстрактному мышлению. При этом человек может не только приобретать, но и целенаправленно передавать другим людям знания, сформулированные в устном или письменном виде в качестве абстрактных понятий, причем практически в неограниченном объеме.
Говоря о предмете зоопсихологии и границах поля деятельности зоопсихолога, следует отметить, что Вагнер считал зоопсихологию лишь частью сравнительной психологии, в которую в качестве второй составной части он включал и психологию человека. В настоящее время сравнительную психологию принято считать наукой о закономерностях происхождения и развития психики животных и человека, об общем и различном в их психической деятельности. Таким образом, предмет сравнительной психологии, выходя за пределы зоопсихологических исследований, включает как обязательный компонент сравнительное (по отношению к животным) изучение психической деятельности человека. Поэтому сравнительно-психологический анализ строится на данных зоопсихологии и психологии человека и направлен на выявление в онто- и филогенезе как сходных психических компонентов, свидетельствующих об общности происхождения психических процессов животных и человека, так и качественных отличий человеческой психики.
Зарождение научной зоопсихологии и сравнительной психологии относится к концу XVIII – началу XIX века, когда появились труды крупнейших биологов того времени – Ж. Л. Л. Бюффона и Ж. Б. Ламарка. Впоследствии особенно большую роль сыграл своими исследованиями Ч. Дарвин, показавший общность происхождения психических процессов животных и человека наравне с общностью происхождения признаков их строения. Однако на начальной стадии развития сравнительной психологии упускались из виду качественные различия психики человека и животного. Широкое распространение в последарвиновский период получили антропоморфические взгляды, а отсутствие научных фактов, как уже отмечалось, восполнялось поверхностными суждениями о психических процессах у животных по аналогии с психической деятельностью человека. Выше уже говорилось о критике Вагнером таких антропоморфических, а также вульгарно-материалистических взглядов на психическую деятельность.
Столь же ошибочными и бесперспективными являются и поныне бытующие, казалось бы, противоположные по своему духу концепции, построенные на полном отрыве психики животных от таковой человека. В этом случае речь идет о субъективно-идеалистическом, антиэволюционном постулате исключительности человеческой психики (что можно объяснить лишь ее божественным сотворением), которая представляется чем-то замкнутым в себе, неким особым «духовным началом», порожденным факторами, не поддающимися естественнонаучному изучению. Сходный взгляд на сущность психического характеризует и «психофизический параллелизм», согласно которому психика существует независимо от материи, подчиняясь лишь своим внутренним «нематериальным» закономерностям. Таким образом, отрицается наличие материальной основы психики и каузальной связи между ней и физиологическими процессами в организме, и постулируются какие-то особые закономерности психической активности, якобы в корне отличные от закономерностей развития органического мира. В итоге закрываются все пути к познанию психического, особенно у животных, где отсутствует возможность самонаблюдения. Но тем самым зоопсихология лишается своего предмета исследования, а следовательно и права на существование.
Отсюда видно, что подлинное изучение психической активности животных возможно лишь с позиции диалектического материализма при понимании психики как продукта развития высших форм органической материи, выполняющей функцию отражения компонентов объективно существующего материального мира. При этом, как уже отмечалось, полноценную и истинную картину психической деятельности можно получить лишь при комплексном изучении поведения и психики животных, учитывая как их единство, так и качественные различия между ними. Поэтому сложившаяся в борьбе с идеалистическими и вульгарно-материалистическими воззрениями современная материалистическая зоопсихология исходит в понимании и изучении психики животных из диалектического единства поведения и психики и строит научный поиск на объективном психологическом анализе структуры поведения животных с учетом экологических и физиологических особенностей изучаемого вида.
При таком подходе снимается и вызвавший недоверие к зоопсихологии как науке довод о мнимой непознаваемости психики животных, о невозможности проникновения в их субъективный мир. Дополнительно к тому, что уже говорилось по этому вопросу, следует отметить, что этот довод воплощает и неверное представление о задачах зоопсихологии. Научная зоопсихология отнюдь не ставит перед собой задачу познания субъективной окраски переживаний животных. Но она может и должна дать ответы, например, на такие вопросы: в чем заключается сущность и характерные особенности восприятий, представлений и других психических категорий у животных, думают ли животные, каковы отличительные признаки их элементарного мышления и т.п.
Наряду с психологическим изучением поведения животных в последние десятилетия получило широкое распространение изучение общебиологических основ и закономерностей поведения животных. Этим занимаются этологи, которых поведение животных интересует прежде всего как экологический фактор, как фактор приспособления животных к условиям среды в ходе индивидуального развития и в процессе эволюции. Одновременно этолог стремится выявить также закономерности изменения поведения в ходе филогенеза и зарождения новых форм поведения. Словом, речь идет о биологических корнях и адаптивном значении поведения. При этом специфика этологического анализа (особенно филогенетических аспектов) и заставляет исследователя обратить внимание прежде всего на видотипичные (генетически фиксированные, т.е. инстинктивные) компоненты поведения, которые проявляют такую же устойчивость и таксономическую (систематическую) достоверность и значимость, как морфологические признаки.
На основе множества полевых наблюдений и экспериментальных данных этологи создали стройную теорию «биологии поведения», в которой центральное место занимает современная концепция инстинктивного поведения. Они внесли ценный вклад в эволюционное учение, в функциональную морфологию, популяционную экологию, систематику животных, зоогеографию, в ряд прикладных отраслей (животноводство, звероводство, охотоведение, рыболовство, охрану природы, бионику) и др.
Итак, зоопсихологи направляют свои усилия на изучение психических аспектов поведения, этологи – биологических. Понятно, что эти два аспекта можно разделить лишь условно: ведь без учета психических факторов невозможно охватить все биологические аспекты поведения. Психика животных не довесок, не эпифеномен, а совершенно необходимый компонент онто- и филогенеза, регулирующий отношения организма со средой. Выдающийся советский зоолог академик А.Н.Северцов убедительно показал, что в процессе эволюции психика играет в определенных ситуациях решающую адаптивную роль. Поэтому развитие психики животных являлось исторической необходимостью, и можно сказать, что без психического отражения действительности, без совершенствования этого отражения не была бы возможна эволюция животного мира.
С другой стороны, зоопсихология не может развиваться без творческого содружества с этологией. Ведь психическое проявляется только в единстве с деятельностью, с внешней активностью, поведением животных. Только опираясь на добытые этологами знания о биологических аспектах и закономерностях поведения животных (особенно относящихся к естественному поведению животных в природных условиях), можно с успехом проникнуть в психический мир животного, который всецело подчиняется биологическим закономерностям и отражает биологические связи организма со средой.
Не менее существенными являются связи зоопсихологии с физиологией, особенно с нейрофизиологией и физиологией высшей нервной деятельности. Но и здесь разделение труда между зоопсихологом и представителем смежной науки выступает достаточно четко, равно как необходимость сплочения их усилий в изучении общего объекта исследования – поведения.
В отличие от зоопсихолога физиолог изучает не само психическое отражение, а обусловливающие его процессы в организме. Физиологическое исследование осуществляется также (как и зоопсихологическое) путем анализа поведения, но под другим углом зрения, с другой целевой установкой. При этом физиолог, естественно, обращает главное внимание на функции нервной системы, головного мозга. Его первейшая задача – изучение деятельности систем и органов, непосредственно или опосредствованно участвующих в поведении животного как целостного организма.
Конечно, при изучении происходящих в организме процессов, обусловливающих психическое отражение, физиолог, подобно зоопсихологу и этологу, должен всегда иметь в виду, что организм представляет собой единое целое. Но поведенческий акт не просто сумма физиологических процессов. На поведенческом уровне, уровне двигательной активности всего организма как единого целого во взаимодействии с комплексом факторов среды, возникает новое качество, определяемое высшим уровнем интеграции всех процессов жизнедеятельности. Сводить поведение к его физиологическим механизмам – значит стирать грань между целым и частичным, между процессами, обусловливающими внешнюю активность организма и самой этой активностью. Поэтому нельзя подходить к этологическим или зоопсихологическим работам с той же меркой, что и к физиологическим. Задачи физиологии иные, чем этологии или зоопсихологии. Ни одна из этих наук не может заменить другую, и только тесное сотрудничество и взаимное обогащение являются условием прогресса каждой из них.
Значение исследования психической деятельности животных для марксистской философии четко определяется замечанием В. И. Ленина в «Философских тетрадях» о том, что «история умственного развития животных» относится к тем областям знания, «из коих должна сложиться теория познания и диалектика».*
* Ленин В.И. Философские тетради. М., 1969. С. 314.
Велико значение данных зоопсихологии и для решения коренных проблем общей психологии, для выявления биологических корней психической деятельности человека и закономерностей происхождения и развития его сознания. Как указывал Выготский, современная психология «рассматривает каждую форму поведения как продукт длительного процесса развития, как форму, исторически связанную с предшествующими более низкими формами».*
* Выготский Л.С. Развитие высших психических функций. С. 398.
Важны знания о психической деятельности животных и для детской психологии, где зоопсихологические исследования помогают выявить биологические основы развития детской психики и – в сравнительно-психологическом плане – ее генетические корни. Для решения чрезвычайно важной и сложной проблемы – развития человеческой психики в онтогенезе – исследователям приходится все обстоятельнее изучать ранние стадии развития, включая и эмбриональный период. А на этих стадиях особенно большую роль играют еще биологические факторы поведения, общие для человека и животных. Так, например, теперь известно, что поведение ребенка, особенно на ранних этапах развития, формируется в большей степени под влиянием «ключевых раздражителей», что существуют «критические периоды», на протяжении которых совершаются процессы «облигатного научения» (преимущественно в форме «запечатления») и т.д. (Эти важные компоненты поведения будут рассмотрены в дальнейшем при характеристике поведения животных.) Вместе с тем зоопсихология вносит свой вклад и в педагогическую психологию, ибо общение детей с животными имеет большое воспитательное и познавательное значение. При таком общении устанавливается сложный психический контакт и взаимодействие между обоими партнерами, что может быть эффективно использовано для умственного и нравственного воспитания детей.
Без данных зоопсихологии, особенно без данных о высших психических функциях животных и особенностях поведения приматов, не может обойтись антропология при решении проблемы происхождения человека. Как будет показано в заключительной главе, эти данные совершенно необходимы для выяснения биологических предпосылок и основ антропогенеза, для изучения предыстории человечества и зарождения трудовой деятельности, в общественной жизни и членораздельной речи.
Зоопсихологические исследования приобретают все большее значение для медицинской практики, где изучение расстройств психической деятельности животных помогает изучать и лечить нервные и психические болезни людей. Большую пользу зоопсихологи приносят сельскому хозяйству, звероводству, рыболовству, охотничьему хозяйству и т.д. Эти исследования приобретают в настоящее время особое значение в связи с далеко идущими преобразованиями в животноводстве и резкими изменениями в среде обитания полезных диких животных. Так, например, при переходе к «промышленному» животноводству существенно ухудшаются условия, необходимые для нормальной психической деятельности сельскохозяйственных животных, что влечет за собой значительное уменьшение их продуктивности (несмотря на хороший уход и полноценное удовлетворение всех их физиологических потребностей). Задачи зоопсихологов заключаются здесь в том, чтобы изыскать возможности компенсации этих неблагоприятных условий и восстановить нормальное поведение животных в столь экстремальных условиях.
Методы зоопсихологических исследований
Как уже указывалось, материалистическая зоопсихология исходит в своем научном поиске из того, что основой и источником психического отражения является у животных их поведение, «животная практика». Качественное различие между последней и практикой человека заключается в том, что животные не поднимаются выше уровня общей приспособительной предметной деятельности, в то время как у человека решающее значение имеет недоступная животным высшая, продуктивная форма предметной деятельности – труд. Вместе с тем психологический анализ конкретных форм двигательной активности животных, структуры их действий, актов их поведения, направленных на отдельные компоненты среды, дает четкое представление о тех или иных психических качествах или процессах.
Конкретно психологический анализ поведения животного осуществляется зоопсихологом путем детального изучения движений подопытного животного в ходе решения определенных задач. Эти задачи ставятся так, чтобы по движениям животного можно было с наибольшей точностью судить об изучаемом психическом качестве. Одновременно должны учитываться физиологическое состояние животного, внешние условия, при которых проводится опыт, и вообще все существенные факторы, способные оказать влияние на результат эксперимента.
Важную роль играют в зоопсихологических исследованиях и наблюдения за поведением животного в естественных условиях. Здесь важно проследить изменения, наступающие в поведении животного при тех или иных изменениях в окружающей среде. Это позволяет судить как о внешних причинах психической деятельности, так и о приспособительных функциях последней. Как в лабораторных, так и в полевых условиях высокоразвитая наблюдательность исследователя является важнейшим залогом успеха его работы.
Хотя изучение структуры поведения животного предполагает в первую очередь качественную оценку его активности, немалое значение имеют и точные количественные оценки в зоопсихологических исследованиях. Это относится к характеристике как поведения животного, так и внешних условий (параметров среды).
Образцом умелого сочетания наблюдения и эксперимента, количественного и качественного анализа поведения животных может служить научное творчество выдающегося советского зоопсихолога Н. Н. Ладыгиной-Котс. Так, например, еще в 1917–1919 гг. она изучала моторные навыки макака с помощью метода «проблемной клетки», т.е. экспериментальной установки, снабженной запирающими механизмами, которые животное должно было отомкнуть. Исследователей, которые до нее пользовались этим методом, по существу, интересовали лишь скорость решения задачи и «потолок» возможностей животного при последовательном усложнении экспериментальной ситуации. Ладыгина-Котс же воспользовалась «проблемной клеткой» с принципиально иной целью – с целью познания психики обезьяны, изучения ее двигательных и познавательных способностей. И поэтому в ходе эксперимента она следила не только за движением стрелки секундомера, но прежде всего за движениями рук подопытного животного, отдавая себе отчет в том, что именно эти движения непосредственно связаны с «душевной жизнью» обезьяны.
Уже в те годы, будучи еще молодым ученым, Ладыгина-Котс искала проявления психического в особенностях двигательной активности животного, в конкретных формах воздействия на окружающие его предметы. И в последующих своих работах она убедительно показывала, что зоопсихолог должен изучать не столько то, что делает животное, сколько то, как оно это делает. Поэтому Ладыгина-Котс предостерегала об опасности ущемления двигательной активности исследуемого животного, ограничения его инициативы и искусственного навязывания определенных движений, так как это неизбежно приводит к искаженным или даже неверным выводам, а одновременно и к утрате ценнейших сведений о психических качествах животного. В связи с этим Ладыгина-Котс всегда относилась с должной осторожностью к результатам изучения психической деятельности животных в одних лишь условиях лабораторного эксперимента, четко видела границы возможностей его применения и дополняла собственные экспериментальные данные результатами наблюдений над свободным, не навязываемым животному поведением.
Весьма важным моментом зоопсихологического исследования является также учет биологической адекватности условий проведения опыта и применяемой методики. Если опыт проводится без учета специфических особенностей биологии изучаемого вида и естественного поведения данного животного в экспериментально имитируемой жизненной ситуации, то результат исследования будет искаженным и легко может оказаться артефактом, что показывает хотя бы следующий пример.
Почти в одно и то же время, в 1913–1914 гг., два выдающихся исследователя поведения животных К.Гесс и К.Фриш изучали способность пчел к различению цветов. Гесс выпускал пчел в темном помещении, где они могли лететь к двум источникам света – разного цвета и различной светлоты. Применив различные сочетания, ученый установил, что пчелы всегда летят к более светлому источнику независимо от длины волны. Отсюда он заключил, что пчелы не различают цвета.
Фриш же, по-иному построив эксперимент, пришел к прямо противоположному выводу. В его опытах пчелам предлагалось на свету выбирать цветные (например, желтые) куски бумаги среди белых, черных и серых различных оттенков, чем уравнивалась интенсивность окраски подкрепляемых цветных и неподкрепляемых ахроматических бумаг. Пчелы безошибочно находили подкрепляемые сиропом желтые (или других цветов) квадратики из бумаги независимо от светлоты и насыщенности их окраски, оставляя без внимания ахроматические листы. Тем самым способность пчел к цветоощущению была доказана.
Ошибка Гесса заключалась в том, что он ставил опыты в биологически неадекватных для пчел условиях – в темноте. В этих условиях не могут проявиться те формы поведения, в которых цветоощущение играет какую-то роль, например при розыске кормовых объектов. Попав днем в темное помещение, пчела будет только искать выход из него. При этом она, естественно, устремится к более светлому отверстию независимо от цвета поступающих через него световых лучей. Таким образом, результаты, полученные Гессом, не могут свидетельствовать о наличии или отсутствии у пчел цветоощущения и поэтому не могут быть использованы для решения поставленного вопроса.
Здесь наглядно обнаруживается тот факт, что реакции животных на одни и те же внешние стимулы могут быть весьма различными в разных жизненных ситуациях и функциональных сферах. В данном примере пчелы в одной ситуации реагируют на цвета, в другой нет. Более того, в одном случае (в сфере пищевого поведения) пчелы реагируют только на цвет, в другом же (в сфере защитного поведения) – только на интенсивность освещения, совершенно игнорируя цветовой компонент. Все это свидетельствует об исключительной сложности экспериментального зоопсихологического исследования и важности создания биологически адекватных условий проведения опытов.
Конкретные методы зоопсихологических экспериментальных исследований отличаются большим разнообразием, хотя все они, как уже говорилось, сводятся к постановке перед животным определенных задач. Приведем лишь несколько основных методов.
Метод «лабиринта». Подопытному животному ставится задача нахождения пути к определенной, непосредственно не воспринимаемой им «цели», которой является чаще всего пищевая приманка, но может быть и убежище («дом») или другие благоприятные условия. При отклонении от правильного пути в отдельных случаях может применяться наказание животного. В простейшем виде лабиринт имеет вид Т-образного коридора или трубки. В этом случае при повороте в одну сторону животное получает награду, при повороте в другую его оставляют без награды или даже наказывают. Более сложные лабиринты слагаются из разных комбинаций Т-образных (или им подобных) элементов и тупиков, заход в которые расценивается как ошибки животного (рис, 1). Результаты прохождения животным лабиринта определяются, как правило, по скорости достижения «цели» и по количеству допущенных ошибок.
Метод «лабиринта» позволяет изучать как вопросы, связанные непосредственно со способностью животных к научению (к выработке двигательных навыков), так и вопросы пространственной ориентации, в частности роль кожно-мышечной и других форм чувствительности, памяти, способности к переносу двигательных навыков в новые условия, к формированию чувственных обобщений и др.
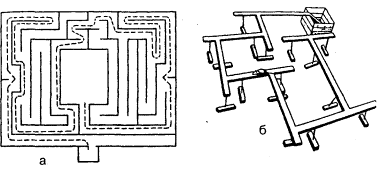
Рис. 1. Лабиринты: а – план первого лабиринта, применявшегося в зоопсихологических исследованиях (лабиринт Смолла); б – лабиринт из «мостиков»
Большинство перечисленных вопросов изучается также методом «обходного пути». В этом случае животному приходится для достижения «цели» обойти одну или несколько преград (рис. 2). В отличие от метода «лабиринта» животное в данном случае непосредственно воспринимает объект (приманку), на который направлены его действия уже в начале опыта. Учитываются и оцениваются скорость и траектория передвижения при поиске обходного пути вокруг преграды. В несколько измененном виде Л. В. Крушинский использовал метод «обходного пути» для изучения способности разных животных к экстраполяции. (Эти эксперименты будут описаны ниже.)
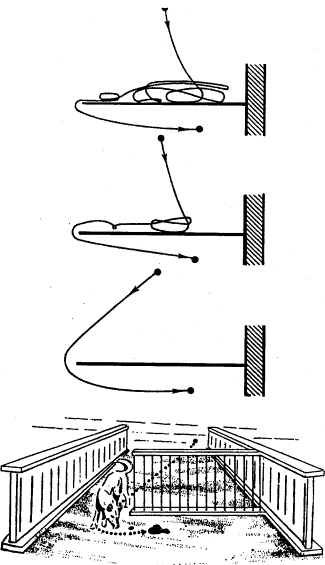
Рис. 2. Постановка опытов по методу «обходного пути» (по Фишелю)
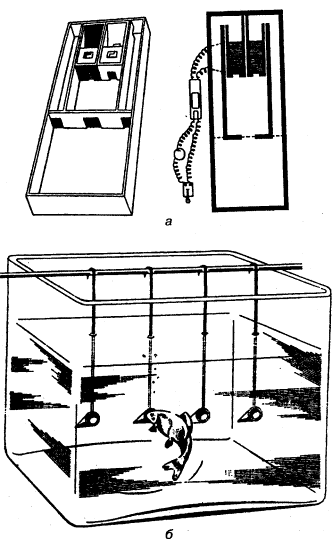
Рис. 3. а – аппарат Йеркса для изучения оптического различения у мелких животных; б – экспериментальное приспособление для изучения цветоразличения у рыб; прикорм находится только в кормушке определенного цвета (опыты Фишеля)
Дифференцировочная дрессировка направлена на выявление способности подопытного животного к различению одновременно или последовательно предъявляемых объектов и их признаков (рис. 3). Выбор животным одного из попарно (или в большем числе) предъявляемых объектов вознаграждается (положительная дрессировка), в других случаях одновременно с подкреплением правильного выбора наказывается неправильный (положительно-отрицательная дрессировка). Последовательно уменьшая различия между признаками объектов (например, их размеры), можно выявить пределы различения (дифференцировки). Таким образом, можно получить сведения, характеризующие, например, особенности зрения у изучаемого вида животных (его остроту, цветоощущение, восприятие величин и форм и т.п.).
Этим же методом изучаются процессы формирования навыков (в частности, на различные сочетания раздражителей), память животных (путем проверки сохранения результатов дрессировки спустя определенный промежуток времени), способность к обобщению. В последнем случае, как правило, постепенно увеличивают несходство последовательно предъявляемых объектов (фигур), выявляя способность животного ориентироваться по отдельным общим признакам этих объектов.
Вариантом дифференцировочной дрессировки, применимым лишь к высшим животным, является метод «выбора на образец». Животному предлагается произвести выбор среди ряда объектов, руководствуясь образцом, который показывается ему непосредственно экспериментатором или в специальном аппарате. Правильный выбор подкрепляется. Этот метод применяется преимущественно также для изучения сенсорной сферы животных.
Метод «проблемной клетки» (ящика). Перед животным ставится задача или открыть для себя выход из клетки, приводя в действие различные приспособления (рычаги, педали, затворы и т.п.), или же, наоборот, проникнуть в клетку, где находится подкорм, отмыкая запирающие устройства. Иногда применяются и небольшие ящики или шкатулки с затворами, отмыкание которых дает подопытному животному доступ к корму. При более сложной постановке эксперимента все механизмы и устройства действуют лишь в строго определенной последовательности, которая должна усваиваться и запоминаться животным. Этим методом исследуются сложные формы научения и моторные элементы интеллектуального поведения животных. Особенно удобно применять этот метод, естественно, для изучения животных с развитыми хватательными конечностями – крыс, енотов, обезьян и др. Это относится и к постановке экспериментов, в которых животным приходится пользоваться орудиями для достижения прикорма. Эти эксперименты также служат преимущественно для выявления высших психических способностей животных.
Элементы орудийных действий отчетливо выступают уже в опытах с применением привязанной к веревке приманки; животное может овладеть- кормовым объектом, лишь притянув его за веревку к себе. Усложняя ситуацию различными сочетаниями веревок и варьируя их взаимное расположение, можно получить ценные данные не только об эффекторных, но и о сенсорных (зрительных и осязательных) компонентах интеллекта животных.
Чаще всего в качестве орудия в экспериментах применяются палки (простые или составные), с помощью которых животные (обычно обезьяны) могут придвинуть к себе или сбить кормовой объект. Широко применяются в опытах с обезьянами (особенно человекообразными) ящики и другие предметы, из которых они должны соорудить «пирамиды» для доставания высоко подвешенного плода. И в этом случае наибольшее значение имеет анализ структуры предметной деятельности животного в ходе решения задачи.
Наряду с такими более или менее сложными экспериментами большую роль играет в зоопсихологических исследованиях анализ обычного, неподкрепляемого манипулирования различными предметами. Такие исследования позволяют судить об эффекторных способностях животных, их ориентировочно-исследовательской деятельности, игровом поведении, способностях к анализу и синтезу и др., а также проливают свет на предысторию трудовой деятельности человека.
Во всех зоопсихологических исследованиях широко применяется фото- и киносъемка, звукозапись и другие средства фиксации поведения животных. Однако никакие технические средства не могут заменить зоркий глаз исследователя и живой человеческий ум, от которых в первую очередь зависит успех в работе с животными.
Часть I
ОБЩАЯ ХАРАКТЕРИСТИКА ПСИХИЧЕСКОЙ ДЕЯТЕЛЬНОСТИ ЖИВОТНЫХ
Глава 1 ПРОБЛЕМА ИНСТИНКТА И НАУЧЕНИЯ
Ранние представления о психической деятельности животных
Начало познания поведения животных
При изучении любой формы психической деятельности прежде всего встает вопрос о врожденном и индивидуально приобретаемом, об элементах инстинкта и научения в поведении животного. В широком плане это вопрос о внешних и внутренних факторах поведения, о его генетических основах и воплощении видотипичного поведения в индивидуальной жизни особи, о мотивации поведения.
Еще древние мыслители, пытаясь постичь основы и движущие силы поведения животных, задумывались над тем, существует ли у животных какое-то «духовное начало», и если да, то какова его природа. Так, Эпикур и его последователи, особенно Лукреций, отвечали на этот вопрос положительно, но при этом со всей определенностью отстаивали положение о материальности такой «души». При этом Лукреций уже высказывал мысль, что целесообразные действия животных являются результатом своего рода естественного отбора, ибо выживать могут лишь животные, обладающие полезными для них свойствами.
Древние философы-идеалисты исходили из представления о некоем изначальном «мире идей», «мировом разуме», аналогичном понятию Бога в более поздних церковных учениях. Порождением этого универсального «разума» является душа человека и животных, которая, по Сократу, соединившись с телом, подвергается влиянию чувствительности и направляется в своих действиях влечениями и страстями. Платон считал, что все поведение животных определяется влечениями. Отсюда был уже только шаг до представления об инстинктах.
Аристотель, «величайший мыслитель древности», как его называл К. Маркс, был и первым подлинным естествоиспытателем среди философов. Человеку Аристотель приписал бессмертную «разумную душу» – воплощение божественного духа. Душа, по Аристотелю, оживляет тленную материю, тело, но только тело способно к чувственным впечатлениям и влечениям. Поэтому в отличие от человека, наделенного разумом, способностью к познанию и свободной волей, у животных имеется лишь смертная «чувственная душа». Но всем животным с красной кровью и рождающим живых детенышей присущи те же пять чувств, что и человеку. Поведение животных направлено на самосохранение и продолжение рода, мотивируется же оно желаниями и влечениями, ощущениями удовольствия или боли. Наряду с этим поведение животных определяется, по Аристотелю, и разумом, представленным у разных животных в разной степени. Разумных животных Аристотель считал способными к пониманию цели.
Свои суждения Аристотель обосновывал конкретными, причем подчас поразительно точными, наблюдениями (наряду, правда, и с анекдотичными данными). У муравьев, например, он подметил зависимость их активности от внешних факторов (освещения). В отношении ряда видов животных он указал на их способность к научению друг у друга; описал он также ряд случаев звукового общения животных, особенно в период размножения. Пожалуй, впервые именно у Аристотеля мы встречаем ссылки на экспериментальные данные о поведении животных. Так, он указывал на то, что после удаления птенцов от родителей они научаются петь иначе, чем последние, и отсюда выводил заключение, что способность к пению не является «даром природы». Таким образом, уже Аристотель обосновывает представление об индивидуальном приобретении определенных компонентов поведения. Эту способность к индивидуальному научению и к запоминанию выученного Аристотель признавал за многими животными и придавал ей большое значение.
Ряд положений Аристотеля получил дальнейшее развитие в учениях стоиков, хотя в некоторых отношениях здесь обнаруживаются и существенные расхождения. У стоиков впервые появляется понятие инстинкта (horme – греч., instinctus – лат.). Инстинкт понимается этими философами как прирожденное, целеустремленное влечение, направляющее движения животного на приятное, полезное и уводящее его от вредного и опасного. Хризипп указывал в этой связи на то, что если утята были выведены даже курицей, их тем не менее влечет к родной стихии – воде, где им обеспечено питание. В качестве других примеров инстинктивного поведения Хризипп ссылался на гнездостроение и заботу о потомстве у птиц, постройку сот у пчел, умение паука ткать паутину. Все эти действия совершаются животными, по Хризиппу, бессознательно, без участия разума (которого у животных нет), без понимания своей деятельности, на основе чисто врожденных знаний. При этом Хризипп отмечал, что подобные действия выполняются всеми животными одного и того же вида одинаково. Таким образом, Хризипп предвосхитил в ряде существенных моментов современный научный взгляд на поведение животных.
Аналогично и Сенека-младший считал, что животные обладают врожденным знанием того, что им вредно, например врожденным знанием внешнего вида хищников. При этом Сенека также указывал на однообразие форм и результатов прирожденной деятельности животных. Четко разграничивая врожденное и благоприобретаемое поведение, Сенека отрицал наличие у животных разума, способности к мышлению. Инстинкт же в концепциях стоиков – властный зов Природы, которому животное вынуждено следовать безоговорочно, «не рассуждая».
Как мы видим, уже в трудах античных мыслителей выступали во всей своей сложности основные проблемы поведения животных: обсуждались вопросы врожденного и благоприобретаемого поведения, инстинкта и научения, а также роль внешних и внутренних факторов психической активности животных. При этом выявлялись те в корне различные подходы к изучению психики животных, которые определяли дальнейший научный поиск в этой области, а именно материалистическое и идеалистическое понимание сущности психической деятельности.
Доэволюционные взгляды на поведение животных (XVIII в.)
После тысячелетнего застоя научной мысли во времена средневековья началось возрождение научного творчества, но только в XVIII в. делаются первые попытки изучения поведения животных на прочном фундаменте достоверных фактов, получаемых в результате тщательных наблюдений и экспериментов. В середине и конце этого века появляются труды целой плеяды выдающихся ученых, философов и натуралистов, оказавших большое влияние на дальнейшее изучение психической деятельности животных.
Из них необходимо прежде всего упомянуть французского философа-материалиста, врача по образованию Ж.- О. Ламеттри. Ламеттри придерживался взгляда, что инстинкты животных представляют собой совокупность движений, выполняемых вынужденно, вне зависимости от размышлений и опыта. При этом он подчеркивал биологическую приспособленность инстинктов, хотя и не мог определить причину этого. Исключительно важной чертой исследований Ламеттри являлся сравнительный подход к изучению психической деятельности животных. Сопоставляя психические способности разных млекопитающих, а также птиц, рыб, насекомых, он показал прогрессивное усложнение этих способностей по направлению к человеку. Осталось сделать только шаг к представлению об историческом развитии психики. Взгляды Ламеттри, сформулированные им на основе тогдашних знаний об анатомии и физиологии нервной системы, оказали впоследствии большое влияние на научное творчество Ламарка.
Крупный французский просветитель Э.Б.Кондильяк в своем «Трактате о животных» (1755) специально рассматривал вопрос о происхождении инстинктов животных. Исходя из сходства инстинктивных действий с действиями, выполняемыми по привычке, Кондильяк пришел к выводу, что инстинкты произошли из разумных действий путем постепенного выключения сознания: разумное поведение превращалось в привычку, а последняя – в инстинкт.
Против такой трактовки категорически возражал Ш. Ж. Леруа. Этот натуралист и мыслитель доказывал, что обозначенный Кондильяком ряд следует читать как бы в обратном порядке. В опубликованных им в 1781 г. «Философских письмах об уме и способности животных к совершенствованию» он выдвигает задачу изучения происхождения разума от инстинкта животных в результате повторяющегося действия ощущения и упражнения памяти. Эту идущую вразрез с церковными догмами концепцию развития высших психических способностей Леруа пытался обосновать и собственными данными о поведении животных в природе. Леруа придавал особое значение полевым исследованиям и настойчиво доказывал, что психическая деятельность животных и особенно их инстинкты могут быть познаны лишь при наличии всесторонних знаний об их естественном поведении и с учетом их образа жизни.
Леруа видел в инстинктах животных воплощение их потребностей: необходимость удовлетворения последних и приводит к появлению инстинктов. Привычки, по Леруа, могут передаваться по наследству и в результате включаться в естественный поведенческий комплекс. Леруа иллюстрировал это на примерах охотничьих собак, передающих свои повадки потомству, или кроликов, перестающих рыть норки после того, как несколько поколений их жило в домашних условиях.
Выше уже упоминалась исключительная роль Ж. Л. .Л. Бюффона в формировании зоопсихологии как науки. Бюффон не только подошел к изучению психической деятельности животных как истинный натуралист, но и сумел при анализе своих полевых наблюдений и опытов воздержаться от антропоморфных толкований поведения животных (в отличие от многих ученых конца XIX и начала XX в.). Согласно его учению, животным (конкретно имелись в виду млекопитающие) свойственны разные формы психической деятельности, в частности ощущения и привычки, но не понимание смысла своих действий. Равным образом животные способны общаться, но их язык выражает лишь чувственные переживания.
Выявляя зависимость между особенностями психической деятельности животных и их образом жизни, Бюффон выдвинул тезис о связи воздействий окружающей среды с внутренним состоянием («желаниями») животного, видя в этом определяющий фактор его поведения. Примечательными являются и его выводы о приспособительном значении психики, а именно о том, что психические качества животных являются не менее значимыми для сохранения видов, чем физические качества. Все концепции Бюффона, построенные на реальных фактах, вошли в созданную им единую систему естествознания и стали первой основой будущей науки о поведении и психике животных. Нам здесь важно отметить, что подлинно научное изучение инстинктов животных (начатое Бюффоном раньше Леруа) привело его к заключению, что сложные действия животных являются результатом сочетания врожденных природных функций, доставляющих животному удовольствие, и привычек. Тем самым Бюффон предвосхитил современное понимание структуры поведения животных, намного опередив своих современников.
Проблема инстинкта и научения в свете эволюционного учения
Ж. Б. Ламарк
К началу XIX в. проблема инстинкта и связанный с ней вопрос о взаимоотношениях врожденных и приобретаемых действий животных привлекают к себе все большее внимание.
Интерес к этим вопросам был обусловлен зарождением идеи трансформизма, появлением первых эволюционных теорий. Актуальной задачей стало выявление того, что передается в поведении по наследству в «готовом виде», что формируется в результате воздействия среды, что является всеобщим, видовым, а что – индивидуально приобретаемым, каково значение разных компонентов поведения в процессе эволюции, где проходит грань между человеком и животным и т.д.
Согласно метафизическим взглядам на природу, господствовавшим на протяжении столетий, инстинкты животных представлялись незыблемыми, пребывающими в неизменном состоянии с момента их «сотворения». Творцам эволюционных теорий предстояло объяснить происхождение инстинктов и на конкретных примерах показать их изменчивость, как они это делали и в отношении видотипичного строения животного организма.
Первое целостное учение об эволюции живой природы было сформулированно в самом начале XIX в. Жаном Батистом Ламарком. Как известно, Ламарк положил в основу своей эволюционной концепции представление о направляющем действии психического фактора. Он полагал, что внешняя среда действует на животный организм опосредованно, путем изменения-поведения животного. Вследствие этого опосредованного влияния возникают новые потребности, которые в свою очередь влекут за собой изменения в строении организма путем большего упражнения одних и неупражнения других органов, т.е. опять-таки через поведение. При всей ошибочности общих положений этой концепции (примат психики как некоего изначального организующего фактора, стремление организмов к «совершенствованию» и т.д.) великой заслугой Ламарка остается то, что он указал на огромную роль поведения, психической деятельности в процессе эволюции.
Стремясь к объяснению всего исторического развития органического мира естественнонаучным путем, Ламарк доказывал, что даже самые сложные проявления психической деятельности развились из более простых и должны изучаться именно в сравнительно-эволюционном плане. При этом он со всей определенностью отрицал существование какого-то особого духовного начала, не связанного с физическим строением животного (как и человека) и не поддающегося естественнонаучному изучению. Все без исключения психические явления Ламарк признавал теснейшим образом связанными с материальными структурами и процессами и поэтому считал их вполне познаваемыми опытным путем, идя по общему пути естественнонаучного познания мира.
Особое значение Ламарк придавал при этом связи психики с нервной системой. В противоположность концепциям психофизического параллелизма, которые бытуют и среди ряда современных этологов, Ламарк убежденно отстаивал положение о функциональной связи психики с нервной системой, о зависимости психической деятельности от нервной деятельности, о зависимости психической эволюции от эволюционных преобразований в нервной системе. Можно с полным основанием сказать, что Ламарк заложил основы сравнительной психологии, сопоставляя строение нервной системы с характером психической деятельности животных на разных уровнях филогенеза.
С этих позиций Ламарк подходил и к проблеме инстинкта. «....Инстинкт животных, – писал он, – это наклонность, влекущая, вызываемая ощущениями на основе возникших в силу их потребностей и понуждающая к выполнению действий без всякого участия мысли, без всякого участия воли».* При этом Ламарк не считал инстинктивное поведение животных чем-то раз и навсегда изначально данным и неизменным. По его представлениям, инстинкты возникли в процессе эволюции в результате длительных воздействий на организм определенных агентов среды. Эти направленные действия и привели к совершенствованию всей организации животного через формирование полезных привычек, которые закрепились в результате многократного повторения, ибо такое повторное выполнение одних и тех же движений приводило к проторению соответствующих нервных путей и все более легкому прохождению по ним соответствующих нервных импульсов («флюидов», по терминологии Ламарка).
* Ламарк. Философия зоологии. Т. 2. М.; Л., 1937. С. 325.
Итак, по мнению Ламарка, тождественная инстинкту наклонность «стоит в связи с организацией, преобразованной привычками в благоприятном для нее направлении, и возбуждается впечатлениями и потребностями, движущими внутреннее чувство особи и предоставляющими ему случай послать – в связи с требованием действующей наклонности – нервный флюид к подлежащим мышцам».* Как отмечал академик В. Л. Комаров, Ламарк понимал под флюидами не что-то мистическое, а вполне материальное начало, близкое к современному пониманию энергии. «...Привычка упражнять тот или другой орган или ту или другую часть тела для удовлетворения вновь и вновь возрождающихся потребностей, – продолжал Ламарк, – делает для нервного флюида... движение в направлении органа, связанного со столь частым употреблением его, настолько легким, что данная привычка становится как бы неотъемлемым природным свойством особи».** Потребности порождают привычки (т.е. обусловливают процесс научения), привычки же превращаются в наклонности, которым животные «не могут противостоять, в которые они сами по себе не в состоянии внести никаких изменений. Отсюда ведут свое начало их привычные действия и их частные наклонности, которым дано название инстинкта».*** Вот как Ламарк представлял себе взаимоотношения между инстинктом и научением: «начало инстинкта в животных» – происхождение инстинктивных действий посредством пожизненного видоизменения двигательной активности.
* Там же.
** Там же.
*** Там же.
Необходимо еще добавить, что, по Ламарку,«наклонность животных к сохранению привычек и к повторению связанных с ними действий... распространяется затем в особях путем воспроизведения, или размножения, сохраняющим организацию и расположение частей в достигнутом состоянии, так что одна и та же наклонность уже существует в новых особях раньше, чем они начали упражнять ее».* Такая передача инстинктов от поколения к поколению «без заметных отклонений будет продолжаться до тех пор, пока не произойдет перемены в условиях, существенных для образа жизни».** В этих словах особенно отчетливо выражено представление об определяющей роли условий среды в эволюции инстинктивного поведения. Конечно, сегодня нам известно, что Ламарк ошибался, полагая, что переход индивидуально приобретенных в наследственно закрепленные формы поведения возможен путем многократного повторения выученных (или видоизмененных) движений, что привычки могут непосредственно обусловить генетические изменения организации животного организма.
* Там же. С. 326.
** Там же. С. 327.
Итак, Ламарк видел в инстинктах животных не проявления какой-то таинственной сверхъестественной силы, таящейся в организме, а естественные реакции последнего на воздействия среды, сформировавшиеся в процессе эволюции. Приспособительный характер инстинктивных действий при этом также является результатом эволюционного процесса, так как постепенно закреплялись именно выгодные организму компоненты индивидуально-изменчивого поведения. С другой стороны, и сами инстинкты рассматривались Ламарком как изменчивые свойства животного. Тем самым взгляды Ламарка выгодно отличаются от встречаемых до наших дней воззрений на инстинкт как на воплощение неких сугубо спонтанных внутренних сил, обладающих изначально целесообразной направленностью действия.
Что же касается индивидуально-изменчивых компонентов поведения животных, их «привычек», навыков, то Ламарк и здесь исходит из материалистических предпосылок, доказывая, что происхождение привычек обусловлено механическими причинами, лежащими вне организма. И хотя Ламарк был не прав, полагая, что сохраненные привычки видоизменяют организацию животного, но в его общем подходе к этой проблеме можно усмотреть правильное понимание ведущей роли функции по отношению к форме, поведения – по отношению к строению организма.
Не давая здесь общей оценки эволюционному учению Ламарка, не касаясь недостатков и исторически обусловленных ошибок этого учения (изначальная целесообразность в природе, в частности в мире животных, гармоничность процесса развития, лишенного противоречий, и др.), необходимо, однако, подчеркнуть трудно переоценимую роль этого великого естествоиспытателя как основоположника материалистического изучения психической деятельности животных и развития психики в процессе эволюции.
К.Ф.Рулье
В середине XIX в. в России страстно и последовательно отстаивал исторический подход к изучению живой природы выдающийся ученый того времени, один из первых эволюционистов, профессор Московского университета К.Ф.Рулье. В те годы в естественных науках все большее распространение получали реакционные теории, и вопросы психики и поведения животных трактовались с идеалистических и метафизических позиций, преимущественно с точки зрения церковного учения. Психическая деятельность животных постулировалась как нечто раз и навсегда данное и неизменное; в их нравах и инстинктах усматривалась беспредельная, непостижимая мудрость Творца, все предусмотревшего и создавшего извечную гармонию в природе.
В этих условиях засилья реакции Рулье решительно и обоснованно выступал против представлений о сверхъестественной природе инстинкта. Подчеркивая, что инстинкты необходимо изучать наравне с анатомией, физиологией и экологией животных, Рулье доказывал, что они являются естественной составной частью жизнедеятельности животных. Это особенно важно, если учесть, что в трудах того времени инстинкты, как и вообще поведение животных, трактовались в отрыве от среды обитания и образа жизни последних. В противоположность упомянутым тео- и телеологическим «объяснениям» Рулье усматривал первопричину происхождения психических качеств и особенностей животных во взаимодействии организма с условиями жизни.
Зарождение и развитие инстинктов Рулье рассматривал как частный случай общей биологической закономерности, как результат материальных процессов, как продукт воздействия внешнего мира на организм. Инстинкт, по Рулье, – это сформировавшаяся на протяжении истории вида, выработанная условиями жизни наследственная реакция на определенные воздействия среды. Конкретные факторы происхождения инстинктов Рулье усматривал в изменчивости, наследственности и постепенном повышении уровня организации животного в ходе исторического развития. В индивидуальной жизни инстинкты могут изменяться в результате накопления опыта.
Иллюстрируя изменчивость инстинктов, Рулье писал, в частности, что подобно тому, как меняются телесные качества животных, «подобно тому, как скот вырождается, как качества легавой собаки, неупражняемой, глохнут, так и потребность отлета для птиц, почему-либо долгое время не отлетавших, может утратиться: домашние гуси и утки сделались оседлыми, между тем как их дикие родичи – постоянно отлетные птицы. Лишь изредка отбившийся от дома домашний селезень во время сиденья его подруги на яйцах, начинает дичать, взлетывать и прибиваться к диким уткам; лишь изредка такой одичавший селезень отлетает осенью с своими родичами в теплую страну и следующею весной опять покажется на дворе, где вывелся».*
* Рулье К.Ф. Избранные биологические произведения. М., 1954. С. 317.
Как перелетный инстинкт, так и нерестовое поведение рыб Рулье пытается проанализировать, опираясь на достижения естествознания его времени. Стремясь к полноценному естественноисторическому объяснению причины перелета птиц, он полагал, что птицы в свое время («когда на нашей планете начали слагаться климаты») начали выбирать себе наиболее благоприятные местности. Молодые птицы «и теперь отлетали бы от нас только в то время, когда голод, холод и нужда принудили бы их к этому, и полетели бы по тому направлению, по которому постепенно уменьшаются невыгодные для их жизни условия...».* Но старые птицы «помнят, как отлетали они прежде... знают, когда наступит срок для заблаговременного отлета... хотя неблагоприятные для них условия еще и не показывались».** Следовательно, в таком сложном явлении, как мифации птиц, Рулье усматривал сочетание и врожденных и благоприобретаемых компонентов, видового и индивидуального опыта. Интересно также отметить, какое значение Рулье придавал при этом передаче опыта от поколения к поколению. Он вообще считал, что у животных «обыкновение, сделавшееся в отце и матери естественною потребностью, отразится и на детях, как отражаются на них вообще привычки и наклонности родителей».*
* Там же. С. 332.
** Там же.
*** Там же. С. 317.
Оценивая зоопсихологические исследования Рулье, необходимо иметь в виду, что в его время (как, впрочем, и впоследствии) термином «инстинкт» часто прикрывали незнание истинных причин поведения животных. Сам Рулье говорил о том, «что инстинкт в большей части случаев – злоупотребляемое словцо, что за ним кроется вопрос, на который наука не нашла еще нужного объяснения...*.
* Там же. С. 441.
Поэтому Рулье стремился при каждом употреблении этого термина наполнить его конкретным содержанием, полученным в результате полевых исследований или экспериментов, делая при этом упор на выявление роли и взаимодействия факторов среды и физиологических процессов.
Именно такой комплексный анализ поведения животных, стремление по возможности полнее выявить определяющие его экологические и исторические факторы и выдвинули труды Рулье на ведущее место среди естествоиспытателей середины прошлого века.
Ч. Дарвин
С победой эволюционного учения Ч. Дарвина в естествознании прочно утверждается мысль о единой закономерности развития в живой природе, о непрерывности органического мира. Сам Дарвин уделял большое внимание вопросам эволюции психической деятельности животных и человека. Им был написан фундаментальный труд «Выражение эмоций у человека и животных», а также ряд специальных работ по поведению животных. Для «Происхождения видов» Дарвин написал специальную главу «Инстинкт». О значении, которое Дарвин придавал изучению инстинктов, свидетельствует уже тот факт, что наличие их у человека и животных как общего свойства он рассматривал как одно из доказательств происхождения человека от животного предка.
Дарвин воздерживался давать развернутое определение инстинкта, но все же указывал, что имеет при этом в виду такой акт животного, который выполняется им «без предварительного опыта или одинаково многими особями, без знания с их стороны цели, с которой он производится».* При этом он с полным основанием отмечал, «что ни одно из этих определений не является общим».**
* Дарвин Ч. Происхождение видов. М., 1952. С. 260.
** Там же.
Ссылаясь на видного исследователя поведения животных первой половины XIX в. Фредерика Кювье (брата знаменитого зоолога и основателя палеонтологии позвоночных Жоржа Кювье), Дарвин сравнивает инстинкт с привычкой и указывает на ряд общих черт их проявления в противоположность сознанию. Но одновременно он предостерегает, что «было бы большой ошибкой думать, что значительное число инстинктов может зародиться из привычки одного поколения и быть наследственно передано последующим поколениям».*
* Там же. С.261.
Происхождение инстинктов Дарвин объяснял преимущественным действием естественного отбора, закрепляющего даже совсем незначительно выгодные виду изменения в поведении животных и накапливающего эти изменения до образования новой формы инстинктивного поведения. Дарвин стремился показать, «что инстинкты изменчивы и что отбор может влиять на них и совершенствовать их».*
* Дарвин Ч. Избранные письма. М., 1950. С. 106.
Свою концепцию Дарвин сформулировал следующим образом: «Едва ли какой бы то ни было из сложных инстинктов может развиться под влиянием естественного отбора иначе, как путем медленного и постепенного накопления слабых, но полезных уклонений. Отсюда, как и в случае с органическими особенностями, мы должны находить в природе не постепенные переходы, путем которых развился каждый сложный инстинкт, – это можно было бы проследить только в ряде прямых предков каждого вида, – но должны найти некоторые указания на эту постепенность в боковых линиях потомков или, по крайней мере, мы должны доказать, что некоторая постепенность возможна, и это мы действительно можем доказать».* В подтверждение того, что происхождение даже весьма сложных форм инстинктивного поведения животных можно объяснить действием естественного отбора, Дарвин подвергает, в частности, обстоятельному анализу гнездовой паразитизм кукушки, «рабовладельческий инстинкт» некоторых видов муравьев и «самый удивительный из всех известных инстинктов – строительный инстинкт пчелы».**
* Дарвин Ч. Происхождение видов. С. 261.
** Там же. С.278.
Действие «одного общего закона, обусловливающего развитие всех органических существ, именно размножения, изменения, переживания наиболее сильных и гибели слабых»,* Дарвин показывает и на многих других примерах поведения диких и домашних животных. Роль наследственности в процессе эволюции инстинктивного поведения он иллюстрирует такими фактами, как, например, обмазывание гнезда глиной у южноамериканских и британских дроздов или замуровывание самок в дуплах у птиц-носорогов Африки и Индии.
* Там же. С.284.
Упражнениям и привычкам, т.е. индивидуальному научению, Дарвин, как уже отмечалось, не придавал сколько-нибудь существенного значения для исторического процесса становления инстинктивного поведения; он ссылался, в частности, на высокоразвитые инстинкты рабочих особей муравьев и пчел, неспособных к размножению и, следовательно, передаче накопленного опыта потомству. «Своеобразные привычки, присущие рабочим или бесплодным самкам, сколько бы времени они ни существовали, конечно, не могли бы воздействовать на самцов и плодущих самок, которые только и дают потомство»*, – писал Дарвин. «И меня удивляет, – продолжал он, – что до сих пор никто не воспользовался этим демонстративным примером бесполых насекомых против хорошо известного учения об унаследованных привычках, защищаемого Ламарком». Дарвин допускал возможность того, что лишь «в некоторых случаях свое влияние оказывают также и привычки и упражнение или неупражнение органа».**
* Там же. С. 283.
** Там же.
Дарвин был не прав, когда признавал единственным критерием развития природы лейбницкое «Natura non facit saltum» («Природа не делает скачков»), приложив этот принцип к инстинктам в равной мере, что и к строению организмов. Но, отстаивая идею взаимозависимости процессов в живой природе и доказывая их материальную сущность, Дарвин показал, что психическая деятельность животных подчиняется тем же естественноисторическим закономерностям, что и все другие проявления их жизнедеятельности.
Очень важным является в этом отношении то, что Дарвин дал обоснованное естественнонаучное объяснение целесообразности инстинктов животных. Как и в отношении признаков строения организма, естественный отбор сохраняет, по Дарвину, полезные изменения врожденного поведения и упраздняет вредные. Эти изменения непосредственно связаны с морфологическими изменениями в нервной системе и в органах чувств, ибо конкретные формы поведения определяются особенностями строения нервной системы, которые передаются по наследству и подвергаются изменчивости, как и все прочие морфологические признаки. Таким образом, целесообразность инстинктов является результатом материального процесса – естественного отбора. Разумеется, это в корне противоречило теологическим воззрениям о сущности психического и его изначальной неизменности, в частности постулату о целесообразности инстинктов как проявлений божественной мудрости.
Дарвин придерживался мнения, «что между развитием умственных способностей и инстинктов существует известное взаимодействие и что развитие последних предполагает некоторое унаследованное видоизменение мозга».* Прогресс умственных способностей, по Дарвину, обусловливался тем, что отдельные части мозга мало-помалу утратили способность отвечать на ощущения «определенным, однообразным, т.е. инстинктивным образом».** При этом Дарвин полагал, что инстинктивные компоненты тем больше господствуют у животных, чем ниже филогенетический ранг последних.
* Дарвин Ч. Происхождение человека и половой отбор. Спб., 1896. С. 45.
** Там же.
Сегодня, спустя сто с лишним лет после этих высказываний Дарвина, мы не можем согласиться с таким противопоставлением основных категорий психической деятельности. Само разделение последней на «однообразно» выполняемые и изменчивые компоненты является условным, поскольку в каждом реальном поведенческом акте ригидные и лабильные элементы поведения выступают в едином комплексе. Соответственно на каждом филогенетическом уровне эти элементы, как будет еще показано, достигнут одинаковой степени развития.
Современное понимание проблемы инстинкта и научения
В проблеме инстинкта и научения большое место занимает вопрос о пластичности инстинктивного поведения. Этот вопрос является весьма важным для уяснения не только эволюции инстинктивного поведения, но и вообще всех вопросов, касающихся психической деятельности животных.
Дарвин считал, что, по существу, одна пластичность инстинктов, проистекающая из изменчивости их врожденных морфологических основ и дающая «материал» для действия естественного отбора, является достаточной для эволюции инстинктивного поведения, а тем самым и поведения вообще. Впоследствии многие ученые посвятили свои усилия изучению того, насколько врожденное, видотипичное поведение является устойчивым или вариабельным, насколько инстинкты постоянны, ригидны или же изменчивы и могут модифицироваться. В результате сегодня мы знаем, что пластичность поведения животных является значительно более сложным явлением, чем это представлялось во времена Дарвина, ибо генетически фиксированы и передаются по наследству не отдельные готовые движения или их сочетания, а нормы реагирования, в пределах которых формируются двигательные реакции в онтогенезе.
Глубокую разработку проблемы инстинкта и научения, как отмечалось, дал В.А. Вагнер, особенно в своем фундаментальном труде «Биологические основания сравнительной психологии» (1910–1913). Опираясь на большой фактический материал, полученный им в полевых наблюдениях и экспериментах и охватывающий как беспозвоночных, так и позвоночных, Вагнер пришел к выводу, что инстинктивные компоненты поведения животных возникли и развились под диктовку среды и под контролем естественного отбора и что их никак нельзя считать неизменными, стереотипными. Инстинктивное поведение, по Вагнеру, – это развивающаяся пластическая деятельность, изменяемая внешними воздействиями.
Вариабельность инстинктивного поведения особенно убедительно была показана Вагнером на примерах конструктивной деятельности пауков и ласточек (рис. 4). Обстоятельный анализ этих фактов привел его к выводу, что лабильность инстинктивного поведения ограничена четкими видотипичными рамками, что стабильными в пределах вида являются не сами инстинктивные действия, а пределы амплитуд их изменчивости. Тем самым Вагнер предвосхитил одно из основных положений современной этологии.
Рис. 4. Гнездостроение у городской ласточки (по Вагнеру). Верхний ряд – контуры разных форм ласточкиных гнезд. Постройки различаются, в частности, относительными размерами крыши (АВ) и дна (CD) гнезда (вид сбоку). Abc – леток. Внизу – схема Вагнера, изображающая вариабельность гнездостроительного поведения. Внутренняя дуга означает среднюю форму и величину летка, внешняя – гнезда. Пунктирные линии – отклонения по длине крыши и боковой стенке. Крайние точки (В1, B2, С1, С2; b1, b2 с1, c2) указывают на видотипичные пределы амплитуд изменчивости этих параметров при нормальном гнездостроении
Впоследствии и другие советские ученые разрабатывали вопросы изменчивости инстинктивного поведения и его связи с процессами научения. Академик Л. А/ Орбели проанализировал зависимость пластичности поведения животных от степени их зрелорождения (см. ч. II, гл. 3). Советский орнитолог А. Н. Промптов указывал на то, что инстинктивные действия животных (птиц и млекопитающих) всегда включают в себя неотъемлемые, очень трудно отчленяемые, но чрезвычайно существенные условнорефлекторные компоненты, формирующиеся в процессе онтогенеза. Именно эти компоненты, по Промптову, обусловливают пластичность инстинктивного поведения. С другой стороны, взаимодействие врожденных реакций, детерминированных видотипичными признаками строения, с приобретенными на их основе в течение индивидуальной жизни условными рефлексами дает в итоге видотипичные особенности, названные Промптовым «видовым стереотипом поведения».
Е. В. Лукина иллюстрировала эти положения Промптова примерами по пластичности гнездостроительной деятельности воробьиных птиц. Так, молодые самки, гнездящиеся впервые в жизни, вьют гнезда, характерные для своего вида. Однако в необычных условиях этот стереотип заметно нарушается. Так, горихвостка и синица-пухляк, являющиеся дуплогнездниками, устраивают при отсутствии дуплистых деревьев свои гнезда под корнями, а серая мухоловка, вьющая гнезда в укрытиях (расщелинах пней, углубленных стволов, за отставшей корой и т.п.), может в случае необходимости устраивать их и на горизонтальных ветках или даже прямо на земле и т.д.
Как мы видим, все это случаи модификации гнездостроительного инстинкта, конкретно – в отношении местоположения гнезда. Описано также немало примеров замены гнездостроительного материала: вместо травинок, мха, лишайника подчас используются такие искусственные материалы, как вата, упаковочные стружки, марля, веревки и т.п. Известны даже случаи, когда мухоловки-пеструшки сооружали свои гнезда в парках Москвы почти целиком из трамвайных билетов. Аналогичные данные были получены и в специальных опытах, в которых исследовалась пластичность инстинктивного поведения при замене яиц или птенцов (опыты Промптева, Лукиной, Скребицкого, Вилке).
У насекомых пластичность строительной деятельности (сооружение чехликов у гусениц Psyche viciella) была в 60-х годах обстоятельно изучена в лаборатории польского зоопсихолога Р. И. Войтусяка (исследования К. Громыша). У другого насекомого, гусеницы Autispila stachjanella, там же изучалась пластичность инстинктивного поведения при устройстве ходов в листьях и коконах (исследования М.Берестынской-Вильчек). Ученые обнаружили большую адаптивную вариабельность инстинктивных действий, особенно при починке сооружений этих насекомых, причем в искусственных, экспериментальных условиях эти действия могут заметно отличаться от выполняемых в обычных, естественных условиях.
Промптов был, безусловно, прав, когда подчеркивал значение слияния врожденных и приобретаемых компонентов во всех формах поведения. Вместе с тем его понимание пластичности инстинктов представляет собой шаг назад по сравнению с концепцией Вагнера, доказавшего, что врожденными являются не инстинктивные действия, а те рамки, в пределах которых эти действия могут выполняться в измененном виде в соответствии с данными условиями среды. Факты, приводимые Промптовым, лишь подтверждают это.
Отказав инстинктивным компонентам поведения во всякой собственной вариабельности, Промптов считал, что пластичность обеспечивается лишь условнорефлекторными компонентами поведенческого акта. В действительности же мы имеем здесь дело с разными по своей величине и своему значению категориями изменчивости поведения. Во-первых, это изменчивость самих врожденных, т.е. инстинктивных, компонентов, которая проявляется в индивидуальной вариабельности видотипичного поведения (в пределах наследственно закрепленной нормы видотипичного реагирования). Примерами такой вариабельности могут служить приведенные Вагнером факты по гнездостроительной деятельности ласточек и упомянутые исследования инстинктивного поведения гусениц. Во-вторых, правда реже, в экстремальных условиях видотипичное, инстинктивное поведение может довольно сильно видоизменяться. Здесь проявляется роль индивидуального опыта, так как именно он обеспечивает более или менее выраженную приспособляемость к необычным, выходящим за рамки нормы внешним условиям. И, наконец, существуют различные индивидуально приобретаемые и поэтому максимально вариабельные формы поведения, в которых главенствующую роль играют уже разные формы научения, имеющие под собой, конечно, тоже инстинктивную основу и переплетающиеся с врожденными компонентами поведения.
Принципиальное значение различий в изменчивости инстинктивного и благоприобретаемого поведения глубоко проанализировал академик А.Н.Северцов, основоположник эволюционной морфологии. В работах «Эволюция и психика» (1922) и «Главные направления эволюционного процесса» (1925) он показал, что у высших животных (млекопитающих) существуют два типа приспособления к изменениям окружающей среды: (1) изменение организации (строения и функций животных), совершающееся весьма медленно и позволяющее приспособиться лишь к очень медленно протекающим постепенным Изменениям среды, и (2) изменение поведения животных без изменения их организации на основе высокой пластичности ненаследственных, индивидуально приобретаемых форм поведения. В последнем случае возможно эффективное приспособление к быстрым изменениям среды именно благодаря изменению поведения. В этом случае наибольший успех будут иметь особи с более развитыми психическими способностями, «изобретатели» новых способов поведения, как метафорически выразился Северцов, – словом, животные, способные выработать наиболее гибкие, пластичные навыки и другие высшие формы индивидуального изменчивого поведения. Именно в этом плане Северцов рассматривает значение прогрессивного развития головного мозга в эволюции позвоночных.
Что же касается инстинктивного поведения, то оно из-за своей малой изменчивости (ригидности) не может выполнять такую функцию. Но подобно изменениям строения тела животного изменения врожденного поведения могут служить приспособлением к медленным, постепенным изменениям среды, поскольку они требуют много времени для своего осуществления.
Напомним, что это изменение врожденного поведения совершается в процессе эволюции на основе индивидуальной изменчивости инстинктивного поведения. Именно в этой индивидуальной лабильности видотипичных действий животных Дарвин искал источник зарождения и развития инстинктов.
Северцов подчеркивал, что значение такого постепенного приспособления не менее важно, чем приспособление с помощью изменения индивидуально приобретаемого поведения. «Инстинкты, – писал он, – суть приспособления видовые, полезные для вида в такой же степени, как и те или другие морфологические признаки, и столь же постоянные».* При этом Северцов обращал внимание на то, что способность к научению, к установлению новых ассоциаций зависит от определенной наследственной высоты психической организации. Сами действия не являются при этом наследственными. В инстинктивном же поведении наследственно закреплено и то и другое. Сегодня мы можем в подтверждение этого тезиса Северцова сказать, что инстинктивное поведение животных определяется врожденной программой действий, реализуемой в ходе накопления индивидуального опыта.
* Северцов А.Н. Собр. соч. Т. III. M.; Л., 1945. С. 300.
Подводя итог изложенному и учитывая современные знания о поведении животных, можно следующим образом охарактеризовать взаимосвязь и взаимозависимость между врожденными и приобретаемыми компонентами поведения и биологическое значение их специфической изменчивости.
Постоянство, ригидность инстинктивных компонентов поведения необходимы для обеспечения сохранности и неуклонного выполнения наиболее жизненно важных функций независимо от случайных, преходящих условий среды, в которых может оказаться тот или иной представитель вида. Во врожденных компонентах поведения хранится итог всего эволюционного пути, пройденного видом. Это квинтэссенция видового опыта, самое ценное, что приобретено в ходе филогенеза для выживания особи и продолжения рода. И эти передаваемые от поколения к поколению обобщенные и генетически фиксированные программы действий не должны и не могут легко изменяться под влиянием случайных, несущественных и непостоянных внешних воздействий. В экстремальных же условиях еще имеются шансы выживания благодаря резервной пластичности инстинктивного поведения в виде модификации.
В остальном реализация врожденной программы поведения в конкретных условиях индивидуального развития животного обеспечивается процессами научения, т.е. индивидуального приспособления врожденного, видотипичного поведения к частным условиям среды. Для этого необходима предельная гибкость поведения, но опять же возможность индивидуального приспособления без утраты существенного, накопленного в ходе эволюции вида, требует незыблемой основы в виде устойчивой инстинктивной диспозиции. Только она дает животному способность с выгодой для себя реагировать в любой ситуации.
Глава 2 ИНСТИНКТИВНОЕ ПОВЕДЕНИЕ
Инстинктивное поведение как основа жизнедеятельности животных
Как было показано, инстинктивное поведение и научение не существуют в реальном поведении сами по себе, а только совместно, переплетаясь друг с другом в единый поведенческий акт. Однако это не означает, что инстинктивное поведение или научение представляет собой лишь условности, искусственно созданные для научного анализа. Условным является здесь лишь их разделение, сами же эти компоненты действительно существуют и функционируют как четко различимые процессы жизнедеятельности со своими специфическими качественными особенностями.
В современных научных исследованиях все чаще избегают употребления понятия «инстинкт» ввиду многообразия и нечеткости его толкования. Понимаемое же как врожденное, наследственно фиксированное, видотипичное «инстинктивное поведение» складывается из инстинктивных действий или актов, которые в свою очередь состоят из отдельных инстинктивных движений (или поз, звуков и т.п.). Четкое разграничение этих терминов необходимо для этологического анализа обсуждаемой категории поведения животных.
Когда мы говорим, что поведение является совокупностью функций внешних, «рабочих» органов животного организма, необходимо различать сами эти функции и их ориентацию во времени и пространстве. И то и другое происходит на инстинктивной основе. Научением же можно только менять ориентацию этих функций. Это означает, что никаким научением нельзя заставить функционировать органы животного иначе, чем это обусловлено их генетически фиксированным строением. Именно морфологические особенности определяют характер функционирования экзосоматических органов, т.е. инстинктивных движений. Нельзя вопреки поговорке научить зайца спички зажигать, так как у него нет соответствующих морфофункциональных предпосылок в строении его конечностей. Но можно научить зайца пользоваться своими конечностями естественным (инстинктивным) образом в нужный момент и в определенном направлении, т.е. ориентировать его инстинктивные движения во времени и пространстве путем научения (в данном случае – дрессировки).
Сказанное не следует понимать в том смысле, что в поведении вообще первичным являются строение, структура, а вторичным – функция, движение. Наоборот, мы говорим о примате движения, функции, имея при этом в виду, что функция определяет форму. Биологическая обусловленность поведения не означает его морфологическую обусловленность; особенно в историческом, филогенетическом плане. В процессе эволюции, несомненно, поведение определяло формирование морфологических признаков, необходимых именно для более успешного выполнения самих поведенческих актов.
Но когда мы говорим о конкретных инстинктивных движениях – результатах эволюционного процесса, то имеем в виду функции именно этих морфологических образований и то, что форма выполнения поведенческих функций обусловливается соответствующими морфологическими структурами. Конкретно это означает, что каждое животное может передвигаться или питаться только так, как это определяется специфическим строением его внешних органов, служащих для выполнения этих функций.
Учитывая все это, можно сказать, что вся жизнедеятельность животного организма, проявляющаяся во внешней активности, зиждется на инстинктивных движениях и других инстинктивных реакциях (термических, электрических, изменения окраски, выделения секретов и т.п.). Они обеспечивают все жизненные функции организма, процессы обмена веществ, а тем самым существование особи и размножение. Вот почему мы говорим о примате инстинктивных движений и по отношению к нервной деятельности, сенсорике, психическому отражению, которые служат у животных лишь для осуществления этих движений, для их ориентации. Поэтому в эволюционном плане развитие психики являлось необходимым следствием (а затем уже – и предпосылкой) повышения уровня обмена веществ и двигательной активности.
Инстинктивное поведение не исчерпывается, однако, самими функциями экзосоматических органов, а включает в себя и механизмы их регулировки и пространственно-временной ориентации. В этом отношении регулировка и ориентация, осуществляемые благоприобретенным путем, на основе научения, служат хотя и важным, но все же лишь дополнением к этим инстинктивным процессам.
Внутренние факторы инстинктивного поведения
Как уже упоминалось, проблема инстинкта и научения непосредственно связана с другой не менее важной проблемой – проблемой внутренних и внешних факторов, мотивации поведения.
Долгое время считали, что инстинктивные действия определяются внутренними, притом загадочными, причинами, в то время как индивидуальное научение зависит от внешних стимулов. В таком виде представления об исключительном или хотя бы преобладающем значении внутренних или внешних факторов встречаются уже у античных мыслителей. При этом мистический, телеологический подход основывался на постулате изначальной целесообразности чисто внутренних, передаваемых от поколения к поколению факторов. Механистический же подход, получивший особое развитие со времен Декарта, признавал лишь внешние факторы движущими силами поведения. В ряде случаев и тот и другой взгляд непримиримо отстаивались вплоть до нашего века.
Что же мы сегодня знаем о мотивации, о движущих силах инстинктивного поведения, а тем самым о поведении вообще? Обратимся сначала к тем внутренним факторам, которые дают первый толчок к любому поведенческому акту, не вдаваясь при этом в детали тех весьма сложных физиологических процессов, которые удалось в этой связи изучить за последние десятилетия.
Внутренняя среда животного организма непрерывно обновляется, но, несмотря на непрекращающиеся процессы ассимиляции и диссимиляции, эта среда остается по своим физиологическим показателям постоянной. Стабильность внутренней среды – совершенно необходимое условие для жизнедеятельности организма. Только при этом условии могут осуществлятся необходимые биохимические и физиологические процессы. Любые, даже незначительные, отклонения от нормы воспринимаются системой интерорецепторов и приводят в действие физиологические механизмы саморегуляции, в результате чего эти нарушения устраняются. Такими механизмами саморегуляции советский физиолог академик П. К. Анохин считал сложные динамические структуры, функционирующие по принципу обратной связи (обратной афферентации) и обозначенные им как функциональные системы (рис. 5).
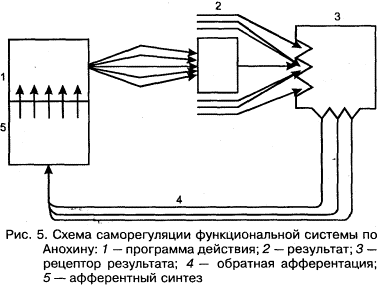
Итак, постоянство внутренней среды основано на самовосстанавливающейся уравновешенности внутренних процессов организма. Важная особенность этих процессов состоит в том, что они протекают в форме ритмов, которые также строятся на системах саморегулирования. Именно в сдвигах этих ритмов видный советский зоопсихолог В. М. Боровский еще в 30-е годы усматривал первичную мотивацию поведения. Выступая против идеалистического понимания инстинктивного поведения, он показал, что в мотивации этого поведения, т.е. в том, что принято называть побуждениями или влечениями, нет ничего сверхъестественного, оторванного от материального мира. Внутреннее побуждение, подчеркивал он, всегда является сдвигом корреляции физиологических ритмов в организме в сторону установления наиболее выгодной в данных условиях коррелированности ритмов всех физиологических процессов. В этом постоянном восстановлении внутреннего равновесия Боровский усматривал основу жизнеспособности организмов.
Итак, первопричиной и основой мотивации поведения являются более или менее значительные и длительные отклонения от нормального уровня физиологических функций, нарушения внутренних ритмов, обеспечивающих жизнедеятельность организма. Эти сдвиги выражаются в появлении потребностей, на удовлетворение которых и направлено поведение.
Первостепенное значение имеют для внутренней мотивации поведения ритмические процессы, происходящие в центральной нервной системе. Собственная ритмика ее стволовой части у позвоночных и брюшных нервных структур у беспозвоночных обеспечивает прежде всего ориентацию поведения во времени. Сейчас хорошо известны автономные, самовозбуждающиеся колебательные процессы («внутренние», или «биологические, часы»), которые регулируют общую ритмику жизнедеятельности организма. В отношении поведения это означает, что периодические колебания внешней активности животных, начало и окончание ритмично повторяющихся действий определяются ритмом «внутренних часов», синхронизированных с космическим временем. Существенные поправки или изменения вносятся в поведенческие ритмы многообразными биологически важными факторами среды, но общая «канва» инстинктивного поведения определяется самовозбуждающимися колебательными процессами с периодом, равным приблизительно суткам (околосуточный, «циркадный» ритм).
В нормальных условиях этот ритм синхронизирован с изменениями в среде, определяемыми вращением Земли вокруг своей оси в течение суток. Однако и в искусственных условиях полной изоляции животного можно наблюдать обычную смену форм активности в те же сроки, что и в нормальных условиях. Это могут быть, например, изменения, связанные со сменой дня и ночи, хотя животное находится в эксперименте в условиях постоянного равномерного освещения.
Помимо циркадных ритмов в поведении животных проявляются и более краткосрочные ритмы, многократно повторяющиеся в течение дня. Так, немецкий этолог В. Шлейдт установил, что клохтанье индюка вполне закономерно повторяется периодически даже в том случае, когда птица полностью изолирована от внешнего мира и даже лишена слуха. Конечно, в нормальных условиях протекание внутренних ритмов изменяется под влиянием внешних воздействий (слуховых, зрительных и прочих стимулов, метеорологических факторов и т.д.), а также зависит от общего физиологического состояния животного.
«Внутренние часы» необходимы и для ориентации животных в пространстве. Хороший пример тому – ориентация птиц во время перелета. Руководствуясь, например, таким астрономическим ориентиром, как солнце, птицы должны учитывать его положение на небосводе в каждое данное время дня, что и происходит путем сопоставления воспринимаемой информации о положении солнца с фазами циркадного ритма.
Уже упомянутый эксперимент Шлейдта показывает, что эндогенные процессы в нервной системе способны обусловливать выполнение определенных инстинктивных движений и при полном отсутствии адекватных внешних стимулов. Так, немецкий нейрофизиолог Э. Хольст обнаружил в стволовой части головного мозга курицы ряд зон, активация которых (в эксперименте – электрическим током) вызывает типичные инстинктивные движения различного функционального значения. При этом оказалось, что при раздражении одного и того же участка мозга, с усилением силы раздражения одно инстинктивное действие сменяется другим в естественной последовательности. Получилась цепь видотипичных движений, выполняемых курицей в определенной биологически значимой ситуации, например при виде приближающегося к ней наземного врага. При этом не только выполнение двигательных реакций, но и последовательность их появления точно так же соответствовала естественному поведению курицы: сперва лишь легкое беспокойство, потом подъем, усиленное беспокойство и, наконец, взлет (рис. 6). Учитывая, что все это происходило в отсутствие каких-либо адекватных внешних раздражителей, становится ясным, что на сугубо эндогенной основе могут выполняться не только отдельные инстинктивные движения, но и целые системы таких движений – инстинктивные действия. Конечно, в естественных условиях такие системы видотипичных, врожденных действий включаются воздействием внешних, экзогенных агентов, в нашем примере – действительным приближением врага, воспринимаемого экстероцепторами. В этом случае постепенное его приближение вызовет усиление раздражения соответствующих участков мозговых структур, что в эксперименте достигалось искусственно при помощи воздействий электрическим током.
Таким образом, поведение в своей основе внутренне так же организовано, так же зиждется на системах биологической саморегуляции, так же закодировано в генетическом фонде вида, как и процессы, определяющие другие функции организма. В этом и проявляется единство всех форм жизнедеятельности животных.
Рис. 6. Последовательное проявление отдельных фаз защитного поведения курицы при продолжительном раздражении одного из участков стволовой части мозга. Длительность и интенсивность раздражения показаны темной полосой (опыты Хольста)
Внешние факторы инстинктивного поведения
Когда говорят об автономности внутренних факторов поведения, об их независимости от внешней среды, то необходимо помнить, что эта независимость является лишь относительной. Уже из приведенных опытов Хольста видно, что эндогенная активность не существует ни «сама по себе», ни «сама для себя»: значение этих спонтанных процессов в центральной нервной системе состоит в прениспозиции к возникновению жизненно важных ситуаций («в случае чего – все готово»). В результате животное способно по первому же сигналу незамедлительно и с максимальной пользой для себя реагировать на изменение в окружающей среде.
Эта готовность обеспечивается тем, что соответствующие эндогенные системы периодически активируются как собственной ритмикой, так и внешними воздействиями (например, изменением продолжительности светового дня, повышением или уменьшением температуры и т.д.). Однако инстинктивные движения, по этологической концепции, заблокированы специальной системой «врожденных пусковых механизмов». Последние представляют собой совокупность нейросенсорных систем, обеспечивающих приуроченность поведенческих актов к биологически адекватным условиям среды (к «пусковой ситуации»). Как только животное оказывается в такой ситуации, соответствующий врожденный пусковой механизм обеспечивает распознавание, оценку и интеграцию специфических для данной инстинктивной реакции раздражителей, после чего наступает растормаживание, снятие «блокировки». Очевидно, одновременно происходит активация соответствующих нервных центров и снижение порогов их раздражимости.
Характерной особенностью для врожденных пусковых механизмов является избирательность реагирования на внешние стимулы: они отзываются только на совершенно определенные комбинации раздражителей, которые только и могут вызвать биологически целесообразную реакцию. Иными словами, в сенсорной сфере существует некая «фильтрующая» функция, выражающаяся в специфической преадаптационной «готовности» воспринимать такие раздражители.
Итак, благодаря врожденным пусковым механизмам внутренняя мотивация поведения получает «выход наружу», т.е. создается возможность без индивидуального опыта в биологически значимых ситуациях реагировать так, чтобы это способствовало сохранению особи и вида.
Подводя итог сказанному, можно сказать, что под врожденным пусковым механизмом следует понимать совокупность нейросенсорных систем, обеспечивающих адекватность поведенческих актов по отношению к «пусковой ситуации»: настройку анализаторов на восприятие специфических раздражителей и распознавание последних, интеграцию соответствующих раздражений и растормаживание (или активацию) нервных центров, связанных с данным поведенческим актом.
Внешние раздражители, составляющие в своей совокупности пусковую ситуацию, получили название «ключевых раздражителей», поскольку они подходят к своим врожденным пусковым механизмам, как ключ к замку. Ключевые раздражители являются такими признаками компонентов среды, на которые животные реагируют независимо от индивидуального опыта врожденными, видотипичными формами поведения, точнее, определенными инстинктивными движениями. В описанном поведении курицы это будут определенные общие признаки, свойственные всем ее наземным врагам.
Кроме собственно растормаживающих ключевых раздражителей (их называют также «пусковыми раздражителями») различают также настраивающие ключевые раздражители, предварительно понижающие порог раздражимости нервных центров, причастных к данным действиям животного, а также направляющие ключевые раздражители, о которых пойдет речь при обсуждении таксисов. Общим свойством всех ключевых раздражителей является то, что это специфические элементарные признаки жизненно важных компонентов среды. Ключевыми раздражителями являются простые физические или химические признаки («просто» форма, размер, подвижность, цвет, запах и т.д.), или их пространственные отношения (взаиморасположение деталей, относительная величина и т.д.), или же векторы. Носителями этих признаков могут быть как другие животные, так и растения и объекты неживой природы. В последнем случае ключевые раздражители выполняют преимущественно направляющую функцию. Так, например, немецкий этолог Ф. Вальтер показал, что у детенышей антилоп ключевым стимулом, определяющим выбор места отдыха (лежа, неподвижно), является «что-то вертикальное» вне зависимости от того, что конкретно это за объект.
Наилучшим образом изучены ключевые раздражители, носителями которых являются животные. Эти раздражители представляют особый интерес и потому, что являются первичными, генетически фиксированными элементами общения у животных. Так, в ставших уже классическими опытах голландского зоолога Н. Тинбергена, одного из основоположников современной этологии, с помощью макетов изучалась пищевая реакция («попрошайничество») птенцов серебристых чаек (клевание клюва родительской особи) и дроздов (вытягивание шеи и раскрытие клюва) при появлении родительской особи.
В естественных условиях голодный птенец серебристой чайки клюет красное пятно на желтом клюве родителя, и тот в ответ отрыгивает пищу в рот птенцу. В опытах предъявлялась серия все более упрощаемых моделей – макетов. Первая модель точно воспроизводила внешний облик естественного носителя ключевых раздражителей, т.е. головы взрослой серебристой чайки с желтым клювом и красным пятном на нем. В последующих моделях путем проб постепенно исключались отдельные детали, и в результате макет становился все менее похожим на голову птицы (рис. 7). В конце концов остался лишь плоский красный предмет с продолговатым выступом. Но этот предмет оказался способным вызвать даже более сильную реакцию птенцов, чем исходная модель. Еще более эта реакция может быть усилена, если этот макет заменить тонкой белой палочкой, исчерченной поперечными темно-красными полосами. Ключевыми раздражителями в данном примере будут просто «красное» и «продолговатое».
Рис. 7. Опыты Тинбергена с применением макетов головы серебристой чайки. Линиями обозначена интенсивность реакций птенцов на макет (число клеваний)
В опытах, проведенных Тинбергеном с десятидневными птенцами дроздов, выявилась другая категория ключевых раздражителей – взаиморасположение и относительная величина деталей объекта-носителя. В качестве макетов использовались плоские диски. Если показать птенцам такой круг, они будут тянуться к его верхней части (местонахождению головы птиц). Если же к большому кругу прибавить маленький, птенцы потянутся к нему. Если, наконец, прибавить к большому кругу два меньших круга, но разных размеров, то решающее значение приобретет относительная величина этих фигур. Размеры «головы» не должны приближаться к размерам «туловища»: при большом «туловище» птенцы потянутся к большему дополнительному кругу («голове») (рис. 8), при маленьком же–к меньшему.
Интересные опыты проводил еще в середине 30-х годов Г.Л. Скребицкий совместно с Т.И. Бибиковой на подмосковном озере Киёво, изучая отношение чайки к ее яйцам. Яйца перекладывались из гнезда в гнездо, заменялись яйцами других видов, искусственными яйцами, а затем и различными посторонними предметами различной величины, формы и окраски. Оказалось, что чайки садились как на чужие, так и на деревянные, стеклянные, каменные, глиняные яйца различной величины и самой разнообразной окраски и начинали их «высиживать». То же самое наблюдалось, когда вместо яиц в гнездо подкладывались разноцветные шары, камешки или картофелины. По свидетельству экспериментаторов, «чайки, сидящие на таких предметах, представляли очень оригинальную картину, но в особенности необычайным становилось зрелище, когда согнанная с гнезда птица возвращалась к нему обратно и, прежде чем сесть, заботливо поправляла клювом разноцветные шары, камешки или картошку».*
· Скребицкий Г. Отношение чайки к гнезду, яйцам и птенцам // Рефлексы, инстинкты и навыки. М., 1936. С. 144–145.
Если, однако, чайкам подкладывались предметы иной формы, например кубики или камни с неровными краями, поведение птиц заметно изменялось. Когда за край гнезда клалось по одному яйцу и инородному предмету, птицы вкатывали обратно в гнездо наряду с яйцами лишь округлые гладкие предметы, размеры которых соответствовали размерам яиц. В известных пределах не препятствовали этому существенные отклонения от нормы в весе (каменное яйцо весило в два с лишним раза больше, чем чаечье), материал, из которого был изготовлен предмет, и окраска.
|
|

Исследователи пришли к выводу, что положительная реакция чайки на яйцо определяется лишь несколькими его элементарными признаками: округлостью, отсутствием выступов, углублений или насечек (рис. 9). Именно эти признаки и выступали здесь как ключевые раздражители.
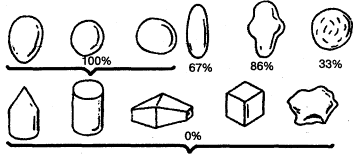
Рис. 9. Вкатывание в гнездо различных предметов в зависимости от их свойств у обыкновенной чайки (в процентах). Верхний ряд, слева направо: яйца (чаечьи, куриные, деревянные и каменные), деревянные шары, другие округлые предметы (картошка, камни) удлиненное глиняное яйцо с углублениями, глиняный шар с зарубками. Нижний ряд, слева направо: деревянный конус, деревянный цилиндр, деревянные и глиняные многоугольники, деревянные и глиняные кубики, угловатые камни (по Скребицкому и Бибиковой)
Важная особенность действия ключевых раздражителей заключается в том, что они подчиняются закону суммации: с увеличением их параметров пропорционально усиливается инстинктивная реакция животного. В экспериментальных условиях это может привести к так называемым «супероптимальным» реакциям, когда животное «преувеличенно», сильнее, чем в норме, реагирует на искусственный раздражитель, в котором «сгущены краски». Мы уже встречались с этим явлением при описании опыта с птенцами чайки, когда они сильнее реагировали на поперечно исчерченную красными полосками палочку, чем на настоящий клюв живой птицы. Суммация проистекает здесь из многократности красных меток и их большей контрастности.
Эффект супероптимальной реакции может в условиях эксперимента привести даже к биологически абсурдному поведению животного. Если, например, предложить чайке два яйца разной величины, она закатит в гнездо более крупное. В итоге может возникнуть такое положение, что птица бросит свое яйцо, чтобы попытаться высиживать деревянный макет яйца гигантских размеров, обладающих супероптимальными признаками ключевого раздражителя.
Как видно из приведенных примеров, ключевые стимулы действуют на поведение животного принудительно, заставляя его выполнять определенные инстинктивные движения, невзирая на возможно воспринимаемую животным общую ситуацию. Это объясняет многие, ранее казавшиеся загадочными моменты в поведении животных.
Так, например, еще в начале нашего века один из авторов антропоморфических сочинений по психологии животных Т. Целл дал следующий ответ на вопрос о том, почему крупные хищники в нормальных условиях при встрече с человеком не нападают на него: например, лев не нападает не потому, что почитает человека, а потому, что не уверен в исходе схватки. «Есть ли у человека оружие, да какое оно, это оружие? – думает лев. – Да ну его, пойду-ка я лучше своей дорогой».
Конечно, лев, как и другие крупные хищники, как правило, уклоняется от нападения на человека не потому, что руководствуется такими глубокомысленными рассуждениями. Разгадка «почтительного» отношения диких зверей к человеку, очевидно, кроется в следующем. Будучи сытым, хищник не реагирует и на присутствие животных, которыми обычно питается. У голодного же хищника преследование животных-жертв и нападение на них обусловливаются сочетанием рассмотренных выше внутренних факторов (первопричина – изменение уравновешенности внутренней среды организма в результате недостатка питательных веществ) с соответствующими внешними стимулами – ключевыми раздражителями, носителями которых являются естественные объекты питания, т.е. животные-жертвы, но не человек.
Даже самый кровожадный хищник не волен нападать на кого угодно и когда угодно. Эти действия также ориентируются во времени и пространстве ключевыми раздражителями, как и все прочие поведенческие акты. Другими словами, дело не в том, «хочет» или «не хочет» животное поступить так или иначе. Если внутреннее состояние животного соответствует определенной внешней пусковой ситуации, то оно волей-неволей вынуждено вести себя так, как это диктует для данных условий генетически зафиксированный код видотипичного поведения.
* * *
Итак, в процессе эволюции возникают адаптации к более постоянным компонентам внешней среды, необходимым для удовлетворения потребностей, непрестанно возникающих в результате изменений внутренней среды организма. Нахождение (или избегание) важных для организма компонентов внешней среды осуществляется путем ориентации по типичным признакам этих компонентов – ключевым раздражителям.
Результаты этой ориентации реализуются нейросенсорными системами (врожденными пусковыми механизмами), которые действуют рефлекторно и включают эндогенные, генетически фиксированные компоненты инстинктивного поведения. Таким образом, действуя «вовне», врожденные пусковые механизмы обеспечивают избирательную направленность внешней активности организма лишь на определенные сигнальные стимулы; действуя же «вовнутрь», они осуществляют оценку и отбор поступающей через рецепторы информации и ее реализацию для активации или понижения порогов раздражимости соответствующих нервных структур, для снятия «блокировки», растормаживания эндогенных нервных процессов, мотивирующих инстинктивные движения и действия. Таким образом осуществляется на врожденной основе корреляция внутренних потребностей организма с биологически существенными изменениями в окружающей его среде.
Только в этой корреляции состоит все биологическое значение эндогенной мотивации поведения. Внутренние стимулы служат лишь для осуществления движения по отношению к среде, без которого организм – именно как саморегулирующаяся система – нежизнеспособен. И в этом, широком, смысле деятельность всей нервной системы в целом всегда рефлекторна.
Равным образом даже наиболее как будто «далеко упрятанные» от внешней среды факторы поведения сами зависят от процессов обмена веществ. Эти процессы уже непосредственно связаны с окружающей организм средой. А так как организм активно регулирует, создает необходимые внешние предпосылки для нормального протекания обменных процессов как раз посредством поведения, то круг смыкается. И в этом отношении проявляется относительность независимости эндогенных автоматизмов, спонтанной активности нервной системы, способности к «самопрограммированию». Ясно, что все эти эндогенные процессы лишь постольку независимы от внешней среды, поскольку они лишь опосредованно связаны с ней.
Структура инстинктивного поведения
Поисковая и завершающая фазы поведенческого акта
Выше говорилось о том, что ключевые раздражители действуют принудительно, что животное вынуждено в своем поведении всецело повиноваться пусковой ситуации. Но значит ли это, что у животных нет никакой возможности проявить собственную инициативу, произвести какой-то самостоятельный выбор? Отнюдь нет!
Инициативное, избирательное отношение животного к среде проявляется прежде всего в активном поиске необходимых пусковых ситуаций и в избрании наиболее эффективных возможностей для выполнения поведенческих актов. Необходимо подчеркнуть, что речь идет о поиске именно раздражений, исходящих от биологически значимых объектов, а не самих этих объектов. Мы теперь уже знаем, что это ключевые раздражители с направляющей или пусковой функцией.
Еще более полувека тому назад американский исследователь поведения животных У. Крэйг показал, что инстинктивные действия состоят из отдельных фаз. Прежде всего Крэйг выделил две фазы, которые в этологической литературе получили названия «поисковая» (или «подготовительная») и «завершающая». В ходе поисковой фазы животное отыскивает (отсюда и название фазы) те ключевые раздражители, точнее, их комбинации (т.е. пусковые ситуации), которые приведут его в итоге к завершающей фазе, в которой воплощается биологическое значение всего инстинктивного действия.
Все промежуточные раздражители не составляют для животного самоцель и ценны лишь постольку, поскольку они приводят к восприятию ключевых раздражителей завершающего поведения. Только на завершающей фазе происходит собственно потребление животным жизненно необходимых ему элементов среды. Но сам поиск адекватных раздражителей является для животных такой же первейшей жизненной необходимостью, как и потребление элементов среды.
Поисковая фаза всегда подразделяется на несколько этапов; однако, в завершающей же фазе, или вовсе не обнаруживаются такие подразделения, или же она состоит лишь из немногих строго последовательно выполняемых движений.
Крэйг построил свою концепцию на данных, полученных им в результате изучения пищевого поведения животных. Приведем пример этой сферы поведения. Хищник, отправляющийся на охоту, вначале еще не знает, где находится его возможная добыча, поэтому первые его движения имеют характер ненаправленного поиска. В результате он раньше или позже попадает в сферу действия стимула, исходящего от животного-жертвы. Обнаружен первый ключевой раздражитель, который включает следующий этап – направленную ориентировку по дополнительным раздражителям, уточнение местонахождения животного-жертвы. Затем следует подкрадывание (или преследование), набрасывание (прыжок) и овладение добычей, ее умерщвление, иногда еще и перетаскивание туши в другое место, расчленение на отдельные куски и, наконец, захватывание кусков мяса зубами и их проглатывание. В этой цепи последовательно выполняемых действий и движений только последние два звена (собственно акт поедания) относятся к завершающей фазе описанного пищедобывательного поведения хищника, все остальные этапы составляют в своей совокупности поисковое (или подготовительное) поведение. Правда, внутри каждого такого этапа имеются свои подготовительные и завершающие фазы, которыми каждый этап и заканчивается. При этом подчас имеется несколько степеней соподчинения (по типу «матрешки»), так что в целом складывается весьма сложная структура деятельности.
Аналогично обстоит дело и в других, казалось бы, значительно более простых сферах поведения, как, например, покой и сон. Животное вначале ищет место для отдыха или ночлега (деревья, укрытия, углубления в почве или просто определенные участки открытого пространства), затем устраивает (совершенствует) найденное место (копает, приминает растительность), иногда еще и чистится и только после этого укладывается (причем видотипичным образом!). Только укладывание составляет завершающую фазу, предшествующие же этапы – поисковую.
К этим примерам можно было бы прибавить еще множество других из любой сферы поведения. Однако уже в приведенных можно усмотреть следующие глубокие различия между обеими фазами, определяющие их сущность.
Поисковое поведение является пластичной фазой инстинктивного поведения. Она характеризуется ярко выраженной ориентировочно-исследовательской деятельностью животных и переплетением врожденных и приобретенных, основанных на индивидуальном опыте компонентов поведения. Именно к поисковому поведению относится все, что связано с пластичностью инстинктов, в частности с модификациями инстинктивного поведения.
Завершающее поведение, наоборот, представляет собой ригидную фазу. Выполняемые в ней движения отличаются строгой последовательностью, стереотипностью и предопределены соответствующими макро- и микроморфологическими структурами. Приобретенные компоненты играют здесь несущественную роль или даже отсутствуют. Поэтому изменчивость ограничена индивидуальной (генетически фиксированной) вариабельностью. Сюда относится все, что говорилось о постоянстве, ригидности инстинктивного поведения и о принудительности действия ключевых раздражителей. Здесь практически все врожденное, генетически фиксированное. Что же касается движений, выполняемых в завершающей фазе, то это и есть собственно инстинктивные движения, или «врожденные двигательные координации», названные так австрийским ученым К. Лоренцем, одним из основателей современной этологии.
Инстинктивные движения и таксисы
Общая характеристика инстинктивных движений уже давалась выше. Также говорилось о том, что они являются «хранителями» самого ценного, жизненно необходимого, что накоплено видом в результате естественного отбора, и что именно этим определяется их независимость от случайных условий окружающей среды. Лисица, производящая инстинктивные движения закапывания мяса на каменном полу, ведет себя «бессмысленно». Но ведь не на каменном субстрате совершался филогенез лисиц, и было бы фатально для дальнейшего существования вида, если бы из-за случайного, временного пребывания особи в таких совершенно нетипичных для местообитания лисиц условиях исчезла бы столь полезная для этих животных форма поведения. Так что лучше произвести закапывающие движения при любых обстоятельствах, и «жесткая» врожденная программа поведения заставляет животное это сделать.
Общая ориентация инстинктивных движений осуществляется таксисами, которые, по Лоренцу, всегда переплетаются с врожденными двигательными координациями и совместно с ними образуют единые инстинктивные реакции (или цепи из нескольких таких реакций).
Как и инстинктивные движения, таксисы – это врожденные, генетически фиксированные реакции на определенные агенты среды. Но если инстинктивные движения возникают в ответ на пусковые раздражители, то таксисы отвечают на направляющие ключевые раздражители, которые неспособны обусловливать начало (или конец) какой-либо инстинктивной реакции, а лишь меняют вектор ее протекания.
Таким образом, таксисы обеспечивают пространственную ориентацию двигательной активности животных в сторону благоприятных или жизненно необходимых условий среды (положительные таксисы) или же, наоборот, от биологически малоценных или опасных условий (отрицательные таксисы). У растений аналогичные реакции выражаются в изменениях направления роста (тропизмы).
По характеру ориентирующих внешних стимулов таксисы подразделяются на фото-, хемо-, термо-, гео-, рео-, анемо-, гидротаксисы (реакции на свет, химические раздражители, температурные градиенты, силу тяжести, течение жидкости, поток воздуха, влажность среды) и т.д. На разных уровнях эволюционного развития таксисы имеют разную степень сложности и выполняют различные функции, о чем еще пойдет речь при обзоре эволюции психики. Сейчас важно подчеркнуть, что таксисы являются постоянными компонентами даже сложных форм поведения, причем высшие формы таксисов выступают в тесном сочетании с индивидуальным опытом животного.
С направляющими ключевыми раздражителями мы уже встречались, когда описывалась ориентация птенцов по относительным оптическим стимулам. Само появление объекта (в эксперименте – диска, в естественных условиях – родительской особи) является пусковым раздражителем реакции «попрошайничества», взаиморасположение деталей этого объекта – направляющим ключевым раздражителем этой реакции, а пространственная ориентация птенцов по этому раздражителю – положительным фототаксисом.
Точно так же красный цвет сам по себе является для птенца серебристой чайки пусковым ключевым раздражителем, обусловливающим его пищевую реакцию (наряду с другим пусковым раздражителем – появлением птицы, точнее, ее головы с клювом). Местонахождение же красного пятна на клюве направляет реакцию птенца биологически выгодным образом на основе положительного фототаксиса и служит, таким образом, направляющим ключевым раздражителем.
Лоренц и Тинберген в 30-е годы совместно изучали взаимоотношения врожденных двигательных координации и таксисов на примере реакции вкатывания яиц в гнездо у серого гуся. Вид яйцеподобного предмета (нечто круглое, без выступов и т.д.), находящегося за пределами гнезда, служит у этой птицы таким же ключевым раздражителем для реакции вкатывания, как у сидящей на гнезде чайки в описанных опытах Скребицкого. Соответствующая врожденная двигательная координация представляет собой многократно повторяемое движение клюва к груди птицы, которое прекратится лишь тогда, когда предмет прикоснется к сидящей в гнезде птице.
Если положить перед краем гнезда перпендикулярно к клюву цилиндр (гуси положительно реагировали и на такой предмет), то подобными инстинктивными движениями и ограничится все поведение птицы. Если же положить яйцо или его макет, то появляются дополнительные движения головы из стороны в сторону, придающие движению предмета правильное направление к гнезду. Ведь в отличие от цилиндра яйцо будет откатываться то влево, то вправо. Вид этих отклонений и служит направляющим раздражителем для таксисных боковых движений головы. Итак, таксисы могут у высших животных ориентировать инстинктивные движения не только всего организма, но и отдельных частей тела и органов.
Из инстинктивных движений и таксисов складывается завершающая фаза каждого поведенческого акта. Вместе с тем они входят как составляющие части в поисковую фазу, которая, как уже говорилось, служит для поиска внешних пусковых ситуаций, позволяющих организму достигнуть завершающей фазы данного акта. Поисковая фаза отличается большой лабильностью и весьма сложной структурой. Инстинктивные движения завершают каждый промежуточный этап этой фазы, в результате конец каждого такого этапа также приобретает черты завершающего поведения. Таксисы дополняются в поисковой фазе ориентировочно-исследовательскими реакциями, непрерывно доставляющими организму информацию о состоянии, параметрах и изменениях компонентов окружающей среды, что позволяет ему произвести оценку последних в рамках общего поискового поведения.
Благоприобретаемые компоненты инстинктивного акта
Наряду с указанными в поисковой фазе любого инстинктивного акта всегда содержатся – в разной степени и в разных сочетаниях – и все элементы поведения, которые относятся к научению, не исключая и высших форм поведения, интеллектуального типа. Именно поэтому мы и считаем, что говорить об инстинктивных действиях (но не движениях!) – значит говорить о поведенческих актах вообще.
В самом деле, все, чему животные способны научиться, направлено лишь к одному – по возможности к скорейшему и наиболее экономному достижению завершающего поведения. Нет в поведении животных ничего, что бы не завершалось этой конечной фазой – инстинктивными движениями или родственными им реакциями, т.е. врожденными двигательными координациями. И в этом отчетливо видно единство инстинктивного поведения и научения.
Конечно, сказанное не относится в равной степени ко всем этапам поисковой фазы. Дело в том, что лабильность поведения неодинакова на разных этапах этой фазы и все больше уменьшается по мере приближения к завершающей фазе.
Переход от одного этапа поискового поведения к другому означает активный поиск и нахождение животными ключевых раздражителей, смену одного этапа поискового поведения другим в строго закономерной последовательности. Это сопровождается ступенчатым сужением сферы активности животного: его поведение все более детерминируется специфическими для данного поведенческого акта сочетаниями раздражителей, становится все более направленным в сторону конечных, завершающих инстинктивных движений, суживаются возможности индивидуального видоизменения поведения животного, пока завершающая пусковая ситуация не сведет эти возможности практически к нулю.
Так, например, ласточка, приступая к постройке гнезда, должна, прежде всего, найти место, где можно собирать гнездостроительный материал. Первоначально ненаправленный осмотр местности является первым этапом гнездостроительной деятельности. Скорость прохождения этого предельно лабильного этапа зависит, прежде всего, от индивидуально-изменчивых компонентов поведения, прежде всего от уже имеющегося индивидуального опыта. В зависимости от своих индивидуальных психических способностей каждая птица решает эту задачу по-своему, более или менее эффективно.
Следующий этап поисковой фазы – поиск и сбор гнездостроительного материала на найденном подходящем месте. Здесь возможности индивидуального изменения видотипичного поведения уже сужены, но тем не менее индивидуальная сноровка продолжает играть немаловажную роль, Еще меньше амплитуда возможных индивидуальных отклонений на основе прежнего опыта на третьем этапе поискового поведения – транспортировке гнездостроительного материала к месту постройки гнезда. Здесь возможны лишь некоторые, не очень существенные вариации в скорости и траектории полета. В остальном поведение всех ласточек уже весьма стереотипное.
И наконец, завершающая фаза – прикрепление частичек к субстрату – выполняется уже совершенно стереотипными инстинктивными движениями. Здесь только генетически обусловленная индивидуальная вариабельность видотипичного поведения дает уже известные нам различия в строении отдельных гнезд.
Таким образом, постепенно уменьшается «свобода действия» животного, уменьшается амплитуда изменчивости его поведения по мере приближения к завершающей фазе в соответствии со все более ограниченными и специфическими условиями среды, сочетаниями раздражителей, причастных к данному поведению.
Чем дальше от завершающей фазы и чем больше амплитуда изменчивости видотипичного поведения, тем больше возможностей включения в него элементов научения, индивидуального опыта и тем больше удельный вес этих элементов. Таким образом, индивидуальный опыт реализуется прежде всего на начальных этапах поискового повелении. И еще: чем выше психическое развитие, тем существеннее коррективы, вносимые в жесткое видотипичное поведение, но опять же главным образом на начальных этапах поисковой фазы. Все это относится, конечно, и к поисковому и завершающему поведению внутри каждого этапа поискового поведения.
Сложность и разнообразие структуры инстинктивного поведения
Двухфазная структура инстинктивных действий дана здесь лишь в виде весьма полной, упрощенной общей схемы. В реальной действительности чаще всего имеют место разные осложнения и видоизменения. Прежде всего необходимо иметь в виду, что поисковая фаза может происходить и под отрицательным знаком – в виде уклонения и избегания определенных агентов среды. Далее возможны сокращения поискового поведения, выпадения отдельных его этапов или даже инверсия. Иногда завершающие движения наступают так быстро, что поисковая фаза не успевает проявиться в полной мере. В других случаях поисковое поведение может свернуть со своего русла и привести к «чужому» завершающему поведению.
Поисковое поведение может принять форму завершающего поведения и существовать наряду с истинной завершающей фазой. В таком случае внешне одинаковые действия будут иметь двоякую качественно различную мотивацию.
Большой интерес представляют и различные случаи неполного протекания инстинктивного акта, когда действия животного не достигают завершающей фазы. У животных с наиболее высокоразвитой психикой промежуточные этапы поискового поведения, т.е. сам по себе поиск раздражителей, могут при этом – как исключение – стать самоцелью их поведения. Здесь мы встречаемся с инстинктивной основой наиболее сложных форм исследовательского поведения, входящих в фундамент интеллекта животных.
Уже этот перечень показывает все разнообразие протекания инстинктивных действий; добавим еще, что практически никогда в одно и то же время не совершается лишь единственный инстинктивный акт, а имеет место сложное взаимодействие между несколькими одновременно происходящими действиями.
Инстинктивное поведение и общение
Все животные периодически вступают во внутривидовые контакты друг с другом. Прежде всего это относится к сфере размножения, где часто наблюдается более или менее тесный контакт между половыми партнерами. Кроме того, представители одного и того же вида часто скапливаются в местах с благоприятными условиями существования (обилие пищи, оптимальные физические параметры среды и пр.). В этих и подобных случаях происходит биологическое взаимодействие между животными организмами, на основе которого в процессе эволюции зародились явления общения. Ни любое контактирование между самцом и самкой, ни тем более скопление животных в благоприятных для них местах (зачастую с образованием колонии) не является проявлением общения. Последнее, как и связанное с ним групповое поведение, предполагает как непременное условие не только физическое или биологическое, но прежде всего психическое взаимодействие (обмен информацией) между особями, выражающееся в согласовании, интегрировании их действий. Как еще будет показано, это относится к животным, стоящим выше кольчатых червей и низших моллюсков.
Дополняя сказанное, необходимо подчеркнуть, что об общении можно говорить лишь тогда, когда существуют особые формы поведения, специальной функцией которых является передача информации от одной особи к другой. Другими словами, общение, в научном значении этого термина, появляется лишь тогда, когда некоторые действия животного приобретают сигнальное значение. Немецкий этолог Г. Темброк, посвятивший много усилии изучению процессов общения и их эволюции, подчеркивает, что о явлениях общения и соответственно подлинных сообществах животных (стадах, стаях, семьях и т.д.) можно говорить лишь тогда, когда имеет место совместная жизнь, при которой несколько самостоятельных особей осуществляют вместе (во времени и пространстве) однородные формы поведения в более чем одной функциональной сфере. Условия такой совместной деятельности могут меняться, иногда она осуществляется при разделении функций между особями.
Добавим еще, что если общение отсутствует у низших беспозвоночных и только в зачаточных формах появляется у некоторых их высших представителей, то, наоборот, оно присуще всем высшим животным (включая и высших беспозвоночных), и можно сказать, что в той или иной степени поведение высших животных в целом осуществляется всегда в условиях общения (хотя бы периодического).
Как уже упоминалось, важнейшим элементом общения является обмен информацией – коммуникация. При этом информативное содержание коммуникативных действий (зоосемантика) может служить опознаванию (принадлежности особи к определенному виду, сообществу, полу и т.п.), сигнализировать о физиологическом состоянии животного (голоде, половом возбуждении и пр.) или же служить оповещению других особей об опасности, нахождении корма, места отдыха и т.д. По механизму действия (зоопрагматика) формы общения различаются каналами передачи информации (оптические, акустические, химические, тактильные и др.), но во всех случаях коммуникации животных представляют собой (в отличие от человека) закрытую систему, т.е. слагаются из ограниченного числа видотипичных сигналов, посылаемых «животным-экспедиентом» и адекватно воспринимаемых «животным-перцепиентом».
Общение между животными невозможно без генетической фиксации способности как к адекватному восприятию (что обеспечивается таксисами), так и к передаче «кодированной» информации (что обеспечивается врожденными пусковыми механизмами). Наследственно закрепленные, инстинктивные действия, с помощью которых выполняется передача информации, могут приобретать самостоятельное значение. Здесь действуют те же закономерности, что и при других формах инстинктивного поведения, но носителями ключевых раздражителей являются в данном случае сородичи или животные других видов.
Среди оптических форм общения важное место занимают выразительные позы и телодвижения, которые состоят в том, что животные весьма заметным образом показывают друг другу определенные части своего тела, зачастую несущие специфические сигнальные признаки (яркие узоры, придатки и т.п. структурные образования). Такая форма сигнализации получила название «демонстрационное поведение». В иных случаях сигнальную функцию выполняют особые движения (всего тела или отдельных его частей) без специального показа особых структурных образований, в других – максимальное увеличение объема или поверхности тела или хотя бы некоторых его участков (посредством его раздувания, расправления складок, взъерошивания перьев или волос и т.п.). Все эти движения всегда выполняются «подчеркнуто», нередко с «преувеличенной» интенсивностью (рис. 10). Но, как правило, у высших животных все движения имеют какое-то сигнальное значение, если они выполняются присутствии другой особи.
Наиболее полноценная, четкая передача информации достигается, однако, тогда, когда появляются специальные двигательные элементы, выделившиеся из «обычных», «утилитарных» форм поведения, утративших в ходе эволюции свою первичную «рабочую», «механическую» функцию и приобретшие чисто сигнальное значение.

Рис. 10. Сигнальные позы и телодвижения у манящего краба (размахивание самцом клешней при «ухаживании» за самкой), рыбы (меченосца), полевого воробья и лошади (по Крейну, Хеменсу, Блуме и Хассенберг)
Первичные движения получили в этологии название «автохтонных» движений; вторичные, приобретшие новую, в данном случае сигнальную, функцию, – «аллохтонных». В таком случае сам рисунок стереотипно, у всех представителей данного вида одинаково выполняемого движения является условным выражением определенной биологической ситуации (биологически значимого изменения в среде или внутреннего состояния животного). Такие видотипичные стереотипные движения с четкой информативной функцией получили в этологии название «ритуализованных движений». Ритуализация характеризуется условностью выполняемых действий, которые служат лишь для передачи определенного, причем совершенно конкретного информативного содержания. Эти действия генетически жестко фиксированы, выполняются с максимальной стереотипностью, т.е., по существу, одинаково всеми особями данного вида, и поэтому относятся к типичным инстинктивным движениям. По этой причине все животные данного вида и в состоянии «правильно» понимать значение подобных оптических (или акустических) сигналов общения. Это типичный пример максимальной ригидности, консерватизма в поведении животных.
Ритуализованные движения выполняются в строгой последовательности в форме более или менее сложных видотипичных «церемониалов» или «ритуалов», причем, как правило, в виде «диалога» двух животных. Чаще всего они встречаются в сферах размножения (брачные игры) и борьбы («мнимая борьба») и отображают внутреннее состояние особи и ее физические и психические качества. Наряду с врожденными существуют и благоприобретаемые формы общения, о которых пойдет речь позже.
Все формы общения играют существенную роль в жизни высших беспозвоночных и позвоночных, обеспечивая согласованность действий особей. Без коммуникации с помощью различных химических, оптических, акустических, тактильных и других сигналов у этих животных невозможно даже сближение и контактирование самцов и самок, а, следовательно, и продолжение рода. Особое значение приобретает общение особей одного, реже разных видов в условиях их совместной, групповой жизни как основа группового поведения.
Как уже говорилось, групповое поведение появляется (по меньшей мере в развитых формах) вместе с общением только у высших беспозвоночных и выражается в согласованных совместных действиях животных, живущих в сообществах. В отличие от скоплений животных, возникающих в результате идентичной положительной реакции на благоприятные внешние условия, сообщества характеризуются относительно постоянным составом его членов и определенной внутренней структурой, регулирующей отношения между ними. Эта структура нередко принимает форму иерархических систем соподчинения всех членов сообщества друг другу, чем обеспечивается сплоченность последнего и его эффективное функционирование как единого целого. Не меньшее значение имеет в этом отношении выделение вожаков, или «лидеров», регулирование отношений между молодыми и старыми животными, совершенствование ухода за потомством и т.д.
Психический компонент инстинктивного поведения
Психический компонент инстинктивного поведения, точнее, психическое отражение при инстинктивных действиях животных необходимо изучать на завершающей фазе поведенческого акта и в какой-то степени на непосредственно примыкающем к ней последнем этапе поисковой фазы, поскольку здесь элементы научения играют наименьшую роль. При этом надо сказать, что в свете современных данных и концепций этологии этот вопрос еще специально не изучался и поэтому приходится здесь ограничиваться лишь некоторыми общими соображениями.
Мы видели, что по мере приближения к завершающей фазе движения животного становятся все более ограниченными, однообразными, стереотипными, и есть все основания полагать, что в такой же мере отражение окружающего мира становится все более бедным, ограниченным. Если на первых этапах поискового поведения животное в поиске ключевых раздражителей руководствуется многими разнообразными ориентирами в окружающей среде, то по мере сужения действия внешних агентов, направляющих поведение животного, ориентация все больше осуществляется только по направляющим ключевым раздражителям, а в завершающей фазе производится уже исключительно по ним.
Вместе с тем мы теперь уже знаем, что как направляющие, так тем более и пусковые ключевые раздражители представляют собой элементарные физические и химические признаки, воспринимаемые животным «в отрыве» от объекта, которому они присущи. Следовательно, животное получает на завершающей фазе лишь весьма неполную, «однобокую» информацию о некоторых – причем несущественных – внешних признаках объектов инстинктивных действий. Правда, эти признаки являются постоянными и в этом отношении характерными для этих объектов.
Однако, что для нас сейчас важно, животное, реагируя на эти признаки, в сущности, не получает сведений о самом объекте (неодушевленном предмете или другом живом существе): признаки – ключевые раздражители – являются только ориентирами, направляющими действия животного на этот объект, носитель этих ориентиров.
Это с совершенной очевидностью вытекает из приведенных выше экспериментов, в которых подлинные объекты инстинктивных действий заменялись весьма абстрактными макетами. Вспомним, что в этих условиях удалось получить даже сверхоптимальные «преувеличенные» инстинктивные реакции. Самец колюшки в завершающей фазе борьбы за гнездовой участок не видит разницы между живой рыбой-соперником и куском жести или пластмассы. Он видит только «красное», и этого ему в данной ситуации вполне достаточно, чтобы среагировать на это «красное», которое в данном случае, кстати, выступает одновременно как направляющий и пусковой ключевой раздражитель.
Следовательно, здесь налицо первично генерализованная реакция на общий элементарный признак (точнее, на совокупность нескольких таких признаков, поскольку в природных условиях действуют не одно только «красное», но и некоторые другие ключевые раздражители). Этот признак воспринимается недифференцированно, как простое ощущение (точнее, воспринимаются суммы ощущений). Об этом свидетельствует отсутствие реакций на форму объекта – носителя признака, да и вообще на объект (предмет или рыбу) как таковой. Ясно, что здесь обнаруживается очень бедное, крайне поверхностное и ограниченное отражение окружающего мира, низшая форма психического отражения. Неинтересно, что такое отражение, свойственное низшим представителям животного мира, в полной мере сохранилось и у высших животных – позвоночных и продолжает у них играть главную роль в поддержании жизнедеятельности особи и продолжении рода.
О том, как это понимать, уже говорилось выше. Эволюция животной жизни могла идти только по пути отбора как можно более простых, но и наиболее постоянных ориентиров (или, что не меняет существа дела, создания таких специфических ориентиров, когда речь идет о животных одного вида, являющихся носителями ключевых признаков), руководствуясь которыми организмы могли бы успешно, не отвлекаясь другими, случайными, изменчивыми признаками, осуществлять наиважнейшие жизненные функции.
Правда, как явствует, например, из описанных опытов с птенцами дроздов, у высших животных ключевые стимулы представлены и более сложными по своей структуре типами. В указанном примере имеет место реакция на конфигурацию и на пространственные соотношения деталей объектов инстинктивных движений, что уже соответствует элементарным восприятиям. Это, очевидно, одно из важных проявлений прогрессивного развития инстинктивного поведения в ходе эволюции. Однако, как показывают те же эксперименты, и здесь нет подлинного предметного восприятия, ибо иначе птенцы не могли бы спутать свою мать с картонным диском.
Примитивность психического отражения на завершающей фазе инстинктивных действий является следствием бедности самой моторики в этой фазе. Как мы знаем, двигательная активность, направленная на окружающую среду, является источником познания этой среды. Однако столь стереотипные движения со столь ограниченной, специальной функцией, какими являются инстинктивные движения, врожденные двигательные координации, не могут служить сколько-нибудь пригодной основой для познания окружающего мира.
Врожденные двигательные координации (во всяком случае в их типичной форме) и не выполняют эту функцию, а ключевые раздражители, воспринимаемые на завершающей фазе (да и на отдельных этапах поискового поведения), не познаются, а узнаются врожденным образом благодаря наличию врожденных пусковых механизмов, чтобы затем использоваться как ориентиры или – и того меньше – как «кнопки» пуска инстинктивных реакций. Здесь больше ничего нет, кроме, вероятно, положительной или отрицательной эмоциональной окраски ощущений от воспринимаемых стимулов и собственных движений.
Познавательные же функции и вообще все богатство психического отражения приурочены к начальным этапам поискового поведения, где в полной мере действуют процессы научения.
Глава 3 НАУЧЕНИЕ
Общая характеристика процесса научения
Анализ инстинктивного поведения приводит к выводу, что все богатство и разнообразие полноценного психического отражения связаны с научением, накоплением индивидуального опыта. Поэтому эта категория поведения будет постоянно в центре нашего внимания и мы еще не раз обратимся к разным ее аспектам, познакомимся с разными формами научения. Здесь же мы рассмотрим лишь некоторые общие положения.
При изучении процессов научения необходимо исходить из того, что формирование поведения представляет собой процесс конкретного воплощения в жизни особи опыта вида, накопленного и закрепленного в процессе эволюции. Именно этим, как уже отмечалось, определяется переплетение врожденных и индивидуально приобретаемых компонентов поведения. Видовой опыт передается из поколения в поколение в генетически фиксированном виде в форме врожденных, инстинктивных компонентов поведения, поэтому «закодированное» таким образом поведение всегда является видотипичным.
Но это не означает, что справедливо и обратное, т.е. что каждая видотипичная форма поведения по своему происхождению обязательно является врожденной, инстинктивной. Существуют такие формы научения, которые внешне очень напоминают инстинктивное поведение, но тем не менее представляют собой не что иное, как результат накопления индивидуального опыта, правда, в жестких видотипичных рамках. Это прежде всего формы так называемого облигатного научения, которым, по Г. Темброку, обозначается индивидуальный опыт, необходимый для выживания всем представителям данного вида независимо от частных условий жизни особи. Относящиеся сюда формы научения будут рассмотрены при обсуждении онтогенеза поведения.
В противоположность облигатному факультативное научение, согласно Темброку, включает в себя все формы сугубо индивидуального приспособления к особенностям тех конкретных условий, в которых живет данная особь. Понятно, что эти условия не могут быть одинаковыми для всех представителей данного вида. Служа, таким образом, максимальной конкретизации видового поведения в частных условиях среды обитания вида, факультативное научение является наиболее гибким, лабильным компонентом поведения животных. О ряде важных аспектов факультативного научения также будет говориться более подробно в связи с онтогенезом поведения (ч. II, гл. 3). Но следует сразу оговорить, что лабильность факультативного научения неодинакова в разных формах его проявления. Конкретизация видового опыта путем вплетения в инстинктивное поведение элементов индивидуального опыта совершается на всех, даже ригидных, этапах поведенческого акта, хотя, как мы уже знаем, в разной степени. Так, английский этолог Рахайнд указывает на модификацию инстинктивного поведения процессами научения путем изменения сочетания раздражителей, обусловливающих определенный инстинктивный акт, выделения (или усиления) определенных направляющих ключевых раздражителей и т.п.
При этом существенно, что модификации могут иметь место как в сенсорной, так и в эффекторной сфере, хотя чаще всего охватывают обе сферы одновременно. В эффекторной сфере включение элементов научения в инстинктивные действия происходит чаще всего в виде перекомбинации врожденных двигательных элементов, но могут возникнуть и новые двигательные координации. Особенно часто это происходит на более ранних этапах онтогенеза. Примером возникновения таких новых двигательных координации может служить подражательное пение птиц. У высших животных, как мы еще увидим, вновь выученные движения эффекторов, особенно конечностей, играют особенно большую роль в познавательной деятельности, в интеллектуальном поведении.
Что же касается «встраивания» индивидуально-изменчивых компонентов в инстинктивные действия в сенсорной сфере, то это существенно расширяет возможности ориентации животного благодаря овладению новыми сигналами. При этом в отличие от ключевых раздражителей, которые являются для животного изначально биологически значимыми, раздражители, которыми животное руководствуется при научении, первоначально являются для него индифферентными (или, точнее, почти таковыми). Лишь по мере того как животное запоминает их в ходе накопления индивидуального опыта, они приобретают для него сигнальное значение. Таким образом, процесс научения характеризуется избирательным вычленением некоторых «биологически нейтральных» свойств компонентов среды, в результате чего эти свойства приобретают биологическую значимость.
Сейчас мы уже знаем, что этот процесс не сводится просто к образованию условных рефлексов. Дело обстоит значительно сложнее, прежде всего благодаря поистине активному избирательному отношению организма к элементам окружающей среды.
Основой этому служат многообразные динамические процессы в центральной нервной системе, преимущественно в высших ее отделах, благодаря которым осуществляется афферентный синтез раздражений, обусловленные внешними и внутренними факторами. Эти раздражения сопоставляются с ранее воспринятой информацией, хранимой в памяти. В результате формируется готовность к выполнению тех или иных ответных действий, что и проявляется в инициативном, избирательном характере поведения животного по отношению к компонентам среды.
Следующий затем анализ результатов действий животного (по принципу обратной связи) служит основой для нового афферентного синтеза. В результате в дополнение к врожденным, видовым программам поведения в центральной нервной системе постоянно формируются новые, благоприобретаемые, индивидуальные программы, на которых зиждутся процессы научения. Физиологические механизмы этого непрерывного программирования поведения были особенно детально проанализированы П. К. Анохиным и получили свое отражение в его концепции «акцептора результатов действия».
Весьма важным является здесь то, что животное не обречено на пассивное «слепое повиновение» воздействиям среды, а само строит свои отношения с компонентами среды и обладает при этом известной «свободой действия», точнее, свободой поиска и выбора.
Итак, формирование эффективных программ предстоящих действий является результатом комплексных процессов сопоставления и оценки внутренних и внешних раздражений, видового и индивидуального опыта, регистрации параметров совершаемых действий и проверки их результатов. Это и составляет основу научения.
Как мы уже знаем, реализация видового опыта в индивидуальном поведении в наибольшей степени нуждается в процессах научения на начальных этапах поискового поведения. Ведь именно на этих первых этапах каждого поведенческого акта поведение животного является наиболее неопределенным, именно здесь от него требуется максимальная индивидуальная ориентация среди разнородных и переменчивых компонентов среды, и особенно важен быстрый выбор наиболее эффективных способов действия, чтобы быстрее и наилучшим образом достигнуть завершающей фазы поведенческого акта. При этом животное может положиться только на собственный опыт, ибо реакции на единичные, случайные признаки каждой конкретной ситуации, в которой может оказаться любой представитель вида, конечно, не могли быть запрограммированы в процессе эволюции.
Но поскольку без включения благоприобретаемых элементов в инстинктивное поведение последнее неосуществимо, само это включение также наследственно закреплено, как и инстинктивные компоненты поведения. Другими словами, диапазон научения также является строго видотипичным. Представитель любого вида не может научиться «чему угодно», а только тому, что способствует продвижению к завершающим фазам видотипичных поведенческих актов. Существуют, следовательно, видотипичные, генетически фиксированные «лимиты» способности к научению.
Рамки диспозиции к научению не только соответствуют реальным условиям жизни, но у высших животных они подчас и шире, чем того требуют эти условия. Поэтому высшие животные обладают значительными резервами лабильности поведения, потенциальными возможностями индивидуального приспособления к экстремальным ситуациям. У низших животных эти возможности резко сужены, как и вообще способность к научению не достигает у них столь высокого уровня, как у высших животных. Поэтому широта диапазонов способности к научению, наличие указанных резервных возможностей являются показателями высоты психического уровня животных. Чем шире эти диапазоны и чем больше потенциальных возможностей к научению сверх обычной нормы, к накоплению и применению индивидуального опыта в необычных условиях, тем больше и лучше может быть скорректировано инстинктивное поведение, тем более лабильной будет поисковая фаза, тем более пластичным будет все поведение животного и тем выше его общий психический уровень.
Это процесс диалектического взаимодействия, при котором причина и следствие постоянно меняются местами: усложнение в процессе эволюции инстинктивного поведения требует расширения диапазона способности к научению; усиленное же включение элементов научения в инстинктивное поведение делает в результате последнее более пластичным, т.е. приводит к качественному обогащению этого поведения, поднимает его на более высокую ступень. В результате поведение эволюционирует как единое целое.
Эти эволюционные преобразования охватывают как содержание врожденных программ поведения, так и возможности их обогащения индивидуальным опытом, научением. На низших этапах эволюции животного мира эти возможности ограничиваются явлениями привыкания, тренировки и суммационными процессами в сенсорной сфере (см. ч. III). Появляющиеся у низших беспозвоночных временные связи еще весьма неустойчивы, затем они становятся все более прочными, достигая полного развития в разного рода навыках. При привыкании постепенно исчезают реакции на определенное многократно повторяющееся раздражение, не сопровождающееся биологически значимым воздействием на животное. Противоположным явлением можно считать тренировку, при которой инстинктивное действие совершенствуется путем накопления индивидуального опыта.
Разница в поведении между низшими и высшими животными состоит не в том, что более примитивные формы поведения сменяются более сложными, а в том, что к первым добавляются вторые, существенно обогащая и изменяя их, в результате чего элементарные формы поведения приобретают значительную сложность и становятся более вариабельным. Так привыкание, с элементарной формой которого мы еще познакомимся на примере поведения простейших, выступает у млекопитающих в весьма сложных проявлениях. Например, как показали опыты, проведенные Хайндом совместно со Скаурсом, на мышах, реакции этих животных на многократно повторяемые, но ничем не подкрепленные акустические стимулы ослабевают с неодинаковой быстротой. Но эти различия в привыкании обусловливаются не только различиями в интенсивности стимулов (как это имеет место у низших животных), но прежде всего вариабельностью самого процесса привыкания.
Основным качественно новым компонентом научения, дополняющим и обогащающим первичные примитивные формы поведения, является навык, который, однако, на разных эволюционных уровнях тоже проявляется в существенно различающихся формах.
Навык
Основные особенности навыков
Навык – центральная, важнейшая форма факультативного научения. Иногда термин «навык» употребляется для обозначения научения вообще. Однако, как подчеркивает А. Н. Леонтьев, нельзя называть навыками любые связи, возникающие в индивидуальном опыте, ибо при таком расширенном понимании понятие навыка становится весьма расплывчатым, а тем самым непригодным для строго научного анализа. Уже указывалось на то, что способность к выработке навыков свойственна не всем животным, а проявляется лишь на определенном уровне филогенеза. В результате формирования навыка применяется врожденная двигательная координация (обычного поведения) в новой сигнальной ситуации или возникает новая (приобретенная) двигательная координация. В последнем случае появляются новые, генетически не фиксированные движения, т.е. животное научается что-то делать по-новому. Но в любом случае решающее значение имеет успешность выполняемых движений и их подкрепление положительным результатом. При этом, по существу, неважно, строится ли научение на информации, полученной путем собственного активного поиска раздражителей, либо в ходе общения с другими животными, коммуникаций, куда относятся также случаи подражания или обучения (человеком – животного, взрослой особью – детеныша и т.п.).
Другой важнейший признак навыка, также связанный с необходимостью подкрепления, заключается в том, что он формируется в результате упражнения и нуждается для своего дальнейшего сохранения в тренировке. При тренировке навыки совершенствуются, при отсутствии же ее угасают, разрушаются.
Навыки интенсивно изучаются у животных с помощью разнообразных специальных методов: «лабиринта», «проблемного ящика» (или клетки), «обходного пути» и др. При всех этих методах животное ставится в условия выбора сигналов или способов действия при решении определенной задачи.
Уже описанный выше метод «лабиринта» был введен в практику экспериментального исследования в 1901 г. американским психологом В. С. Смоллом. В настоящее время применяются лабиринты самой различной конструкции (см. рис. 1), но задача, которую приходится решать подопытному животному, всегда одна и та же – как можно быстрее и без захода в тупики достигнуть места подкрепления (чаще всего пищевого). Большинство опытов проводилось с крысами, которые легко справляются с такими задачами. Практически не существует лабиринта, в котором крыса не сумела бы сориентироваться. Особенно интенсивно пользовались лабиринтами, как и проблемными ящиками и клетками (также уже описанными), бихевиористы. Проблемный ящик был усовершенствован и снабжен автоматическими устройствами американским психологом Б. Ф. Скиннером. В качестве подопытных животных при этом также использовались крысы.
В лабиринте животное решает задачу как бы «вслепую», так как реакция (пробежка) вырабатывается без непосредственного контакта с обусловливающим ее стимулом (пищей, гнездом), и более того, животное в начале опыта еще ничего не знает о наличии целевого объекта, а лишь случайно обнаруживает его впоследствии в результате ориентировочно-исследовательских действий. Запоминание этого объекта и пути к нему и служат основой формирования навыка. Если впоследствии животное многократно пробежит это расстояние одним и тем же выученным кратчайшим путем, то этот навык становится стереотипным, автоматизированным. Пластичность поведения в этом случае невелика, а при сильно выраженной стереотипности выученных движений последние подчас приближаются к инстинктивным двигательным стереотипам. Стереотипность вообще характерна для примитивных навыков, при которых формируются ригидные, автоматизированные двигательные реакции. Значительная пластичность проявляется при таких навыках лишь па первых этапах образования этих навыков. (С навыками высшего порядка, отличающимися большой пластичностью, мы познакомимся позже.)
Выработка навыков по методу Скиннера получила название «оперантное», или «инструментальное», обусловливание. Животное должно при решении таких задач проявить двигательную инициативу, самостоятельно «изобрести» способ действия, оперирования. Животному здесь не «навязываются» экспериментатором определенные движения, как это имеет место при выработке «классических» условных рефлексов по методу Павлова. При обучении крысы в скиннеровском «проблемном ящике» временная связь образуется после ряда случайных движений (нажатий на планочку), подкрепляемых появлением пищи в кормушке. В более сложно устроенных установках крысам предоставляется даже возможность выбора между двумя способами действия, приводящими к разным результатам. Так, например, можно крысе дать возможность «по своему усмотрению» регулировать температуру в «проблемном» ящике, включая то обогревающий рефлектор, то охлаждающий вентилятор в соответствии со своими потребностями. При каждом нажатии на рычаг попеременно включается рефлектор и выключается вентилятор или наоборот (рис. 11).
При «классическом», павловском обусловливании наблюдается «респондентное» поведение, т.е. ответ следует за стимулом, а в результате образования условно-рефлекторной связи и подкрепление (безусловный раздражитель) связывается со стимулом. При оперантном же обусловливании вначале производится движение (ответ), сопровождаемое подкреплением без условного раздражителя. Но как и при «классической» выработке условных рефлексов, адекватная реакция животного (в данном случае двигательная) подкрепляется здесь полезным для животного результатом. При этом потребность в пище или создании оптимальных температурных условий побуждает крысу реагировать на рычаг выученным образом, точнее, ориентировать свое поведение в соответствии с восприятием рычага. Это восприятие действует как условное раздражение постольку, поскольку действие рычагом приводит к биологически значимому результату – пищевому или температурному подкреплению. Вне этой временной связи рычаг не имеет для крысы никакого значения.

Рис. 11. Крыса в скиннеровском «проблемном ящике» (объяснения см. в тексте)
Тот факт, что при павловском обусловливании действует сначала условный агент (условный раздражитель, например звук метронома или вспышка лампочки), а уже затем безусловный (например, вкусовое ощущение в результате пищевого подкрепления), не следует переоценивать, так как и условный рефлекс может быть выработан лишь при наличии внутренней готовности животного, т.е. соответствующей потребности. У вполне сытого животного невозможно выработать условный рефлекс на пищу.
Выше уже говорилось о роли первичной (инстинктивной) мотивации в поведении животных.
По существу, оперантное обусловливание по Скиннеру в сравнении с «классической» павловской постановкой эксперимента охватывает лишь упрощенный, укороченный поведенческий акт: вне внимания оставляется первая фаза – фаза ориентации животного во времени и пространстве. При выработке «классического» условного рефлекса животное должно сперва по внешним ориентирам научиться тому, когда, при каких сопутствующих внешних условиях следует произвести соответствующее движение. Вот эта первичная ориентация животного во времени по внешним сопутствующим признакам не учитывается при упомянутой постановке опытов по методу Скиннера. Равным образом не учитывается и пространственная ориентация, т.е. как животное находит рычаг, как научается пользоваться им, – словом, как ориентирует свои движения в пространстве. А ведь это является самым важным для подлинного психологического анализа поведения животного. Вместо такого анализа структуры деятельности при формировании двигательного навыка бихевиористами учитываются лишь временные параметры этого процесса (например, число нажатий за определенное время) без соответствующего качественного анализа.
В противоположность этому «классическая» павловская методика уже по самому существу своему принципиально верно моделирует естественные условия формирования поведения животного, так как направлена на анализ первичной ориентации животного по признакам компонентов среды, по которым животному необходимо ориентироваться в самом начале любого поведенческого акта. Преимущества такого подхода становятся ясными, если вспомнить, что говорилось выше о первом этапе поискового поведения.
Конечно, как отмечалось, мы сегодня уже не можем при изучении процессов научения, в частности навыков, ограничиваться «классическими» условными рефлексами. И разумеется, «проблемные ящики», лабиринты и тому подобные установки совершенно необходимы для изучения формирования навыков в разных условиях, для выявления способностей животных к решению задач разной степени сложности. Важно только наряду с количественным произвести и качественный анализ структуры поведения животного в ходе решения поставленной перед ним задачи.
Дрессировка
Если при оперантном обусловливании животному предоставляется максимальная возможность проявить инициативу, самостоятельно выбрать способ действия в ходе решения задач, то при дрессировке, наоборот, навыки вырабатываются всецело под целенаправленным воздействием человека и в соответствии с его замыслом. Дрессировка осуществляется путем систематической тренировки животного, при которой подкрепляются требуемые двигательные реакции и их сочетания и упраздняются нежелательные движения. В результате дрессировки, как правило, вырабатываются прочные и четко скоординированные двигательные акты, достигающие подчас большой сложности. Выполняются они животным всегда в ответ на подаваемые человеком сигналы.
При цирковой дрессировке, рассчитанной на зрелищный эффект, раньше практиковался преимущественно болевой метод – наказание (иногда весьма жестокое) животного за каждое неправильное движение. В настоящее время его в основном сменил гуманный (и более эффективный) метод дрессировки, основанный на учете особенностей естественного поведения животного, его предварительном приручении, хорошем с ним обращении и пищевом подкреплении правильно выполняемых движений. Иногда применяется смешанный метод – поощрение правильных и наказание за неправильные движения. Дрессировка находит также широкое практическое применение во многих сферах хозяйственной и военной деятельности человека (служебное и охотничье собаководство).
В научных исследованиях широко пользуются разными формами дрессировки для изучения различных аспектов психической деятельности животных. Четкость условий, в которые ставится животное, и возможность точного учета сигнальных раздражителей (подаваемых экспериментатором сигналов) делают научную дрессировку незаменимой при изучении навыков животных.
Дрессировка является значительно более сложным процессом, чем простое обусловливание, и не сводится лишь к выработке цепей условных рефлексов. Специфическая трудность заключается в том, чтобы дать животному понять, что от него требуется, какие движения оно должно выполнить. Между тем эти движения хотя и входят в видотипичный репертуар поведения, являются зачастую непривычными или трудноосуществимыми в заданных дрессировщиком условиях. Советский зоопсихолог МА.Герд, детально разработавшая теорию дрессировки, делит поэтому процесс дрессировки на три стадии: наталкивания, отработки и упрочения.
На первой стадии решается задача – вызвать впервые ту систему движений, которая нужна человеку, «натолкнуть» животное на ее выполнение. Это достигается тремя путями: путем непосредственного наталкивания, косвенного наталкивания или сложного. В первом случае дрессировщик заставляет животное следовать или поворачиваться вслед за пищевым или иным привлекательным для животного объектом. Во втором случае провоцируются движения, непосредственно не направленные на приманку, но обусловливаемые общим возбуждением животного (так формируются преимущественно манипуляционные действия, выполняемые конечностями: обхватывание, перенос, удары или царапанье лапами и т.д.). Так, например, для создания циркового номера (раскатывание ковра лисицей) дрессировщик, стоя около свернутого ковра, поддразнивает ее куском мяса, но не дает схватить его. Возбужденное животное подскакивает, привстает, перебирает передними лапами и т.д. Все случайные прикосновения к ковру при этом закрепляются небольшими кусками пищи, в результате чего лисица все чаще будет обращаться к ковру, и наконец, появятся нужные дрессировщику движения лапой по ковру. В дальнейшем эти движения отрабатываются, направляются на середину рулона и т.д.
При сложном наталкивании, по Герд, дрессировщик вначале вырабатывает у животного определенный навык, а затем изменяет ситуацию, заставляя животное по-новому применять выработанное умение. Так, балансирование мячом на кончике носа вырабатывается у морских львов после того, как они научились сбрасывать его в руки дрессировщику. Убирая руки из поля зрения зверя, пряча их за спину, дрессировщик заставляет его несколько задерживать мяч на кончике носа, ибо подкрепление (рыба) будет получено животным лишь после того, как мяч окажется в руках человека. Путем обильного подкрепления постепенно увеличивается длительность удерживания мяча, и в конечном итоге получается знаменитый коронный цирковой номер.
Вторую стадию дрессировки, стадию отработки, Герд определяет как этап, на котором совершается отсечение многих лишних движений, вначале сопровождающих необходимые действия животного; далее – отшлифовка первичной, еще весьма несовершенной системы движения и, наконец, выработка удобной сигнализации, с помощью которой дрессировщик в дальнейшем управляет поведением животного. Усилия дрессировщика на этой стадии направлены на упразднение ориентировочных реакций, движений, обусловленных страхом, и иных помех, а также на упорядочивание последовательности, направленности и длительности вырабатываемых движений. Необходимо также заменить реакцию на пищу реакцией на подаваемый дрессировщиком сигнал. При всем этом вновь используются некоторые приемы наталкивания. Например, чтобы отшлифовать у медведя удерживание бутафорного «торта», используется «дробное» наталкивание: дрессировщик поднимает пищевую приманку на нужную высоту и относит ее чуть вбок, в результате чего коробка, которая раньше прижималась медведем к низу живота, поднимается им на уровень груди и немного в сторону. Это правильное положение фиксируется минимальным или средним подкреплением. Равным образом правильная осанка медведя фиксируется приманкой, удерживаемой над его головой, и т.д. С помощью наталкивающих воздействий; производится и выработка искусственной сигнализации.
Заключительная стадия процесса дрессировки, стадия упрочения, характеризуется усилиями дрессировщика, направленными на закрепление выработанного навыка и надежность его воспроизведения в ответ на подаваемые сигналы. Дробное наталкивание (приманивание) применяется уже крайне редко, а пищевое подкрепление осуществляется; уже не после нужного элемента, навыка, а преимущественно после целого комплекса выполненных движений. Вообще кормление производится реже, на более крупными порциями. Выработанные в результате навыки приобретают стереотипную форму при которой конец одного действия может послужить началу последующего.
Произведенный Герд анализ процесса дрессировки убедительно показывает исключительную сложность, гетерогенность в многоплановость поведения животных при выработке у них искусственный навыков. Но не менее сложная картина обнаруживается и для формировании навыков у животных и в естественных условиях. Об этом всегда следует помнить при ознакомлении с лабораторными экспериментами, где предельное искусственное упрощение и схематизация форм поведения животных являются залогом успеха научного исследования.
Познавательные процессы при формировании навыков
Еще в начале нашего века сложилось мнение, что образование навыков – как в отношении ориентации среди элементов среды, так в отношении формирования новых сочетаний движений – происходит путем «проб и ошибок». К этому выводу пришел в результате своих исследований ряд выдающихся ученых – Г. Спенсер, К. Ллойд-Морган, Г. Дженнингс, и, прежде всего Э. Торндайк. Согласно концепции «проб и ошибок», животное запоминает то, что случайно привело к успеху, все остальное постепенно отсеивается. Иными словами, в результате «проб и ошибок» совершается отбор и закрепление случайно произведенных удачных движений, что .и приводит в конце концов путем многократных повторений к формированию двигательного навыка. Конечно, при этом отсутствует какое бы то ни было понимание связей и отношений между компонентами научения. Существенным здесь является представление, что «пробы и ошибки» совершаются беспорядочно.
Однако, как уже было доказано, образование навыков является значительно более сложным процессом и определяется активным отношением животного к воздействующим на него факторам среды. Еще в 20-е годы Э. Толмен, В. П. Протопопов и другие возражали против представления о хаотичности движений, производимых животными при решении задач, и показали, что эти движения формируются в процессе активной ориентировочной деятельности. Пои этом животное анализирует ситуацию и избирает то направление движений, которое соответствует положению «цели». В результате движения животного становятся все более адекватными ситуации, в которой дана задача. Таким образом, на место случайного возникновения движений ставится, как решающий фактор, активный двигательный анализ ситуации.
Этот взгляд получил подтверждение в ряде экспериментальных исследований. Так, американский ученый И. Ф. Дэшиелл показал, что пробные забеги крыс в тупики лабиринта вовсе не случайны, а, как правило, производятся в сторону «цели»: после первой ориентировки в лабиринте животное создает как бы общую систему направления своего передвижения; при этом крыса значительно чаще заходит в тупики, расположенные по направлению к цели, чем расположенные в обратном направлении (рис. 12). Аналогичные данные были получены К. Спенсом и В. Шипли (рис. 13).
Направленность действия при выработке навыков, возникающая в результате первоначальной активной ориентировки животного, побудила И. Кречевского выдвинуть тезис о появлении у животного своего рода «гипотез», которыми оно руководствуется при решении задач. Особенно это выявляется в тех случаях, если подопытному животному ставится заведомо неразрешимая задача, например, когда в беспорядочной последовательности закрываются и открываются проходы лабиринта (рис. 14). В этом случае у разных групп подопытных животных появляются разные, но всегда устойчивые типы поведения. По Кречевскому, животные пытаются выйти из затруднения, строя «гипотезу» и испытывая ее пригодность. В случае неудачи животное заменяет ее другой «гипотезой». Поэтому действия по одной «гипотезе» повторяются многократно, до выявления ее непригодности. Соответственна животное некоторое время ведет себя одинаковым образом независимо от меняющихся внешних условий. Так, в упомянутом лабиринте Кречевского которые крысы, например, первоначально сворачивали на всех развилках в одну и ту же сторону. Убедившись, что эта «гипотеза» не приводит к успеху, они стали постоянно сворачивать в противоположную сторону.
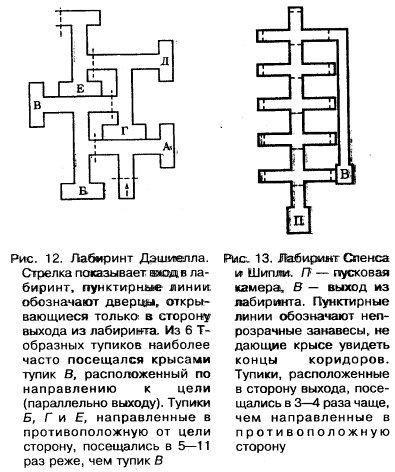
В других случаях крысы начинали регулярно чередовать повороты налево и направо. Таким образом, обнаруживается четкая связь между предыдущей попыткой и последующей, животное как бы стремится организовать свое поведение по одному «принципу». Кречевский считал, что этот в известней мере абстрактный «принцип» отличается систематичностью и обусловлен внутренней «настройкой» животного.
Рис. 14. Лабиринт Кречевского. П – пусковая камера. Прерывистыми линиями показаны дверцы, которыми можно закрывать проход то с левой, то с правой стороны, пунктиром – путь животного. Пояснения см. в тексте
Концепция Кречевского, безусловно, не свободна от недостатков, и сам термин «гипотеза» является крайне неудачным применительно к поведению животных. Вместе с тем большая заслуга этого ученого заключается в том, что он убедительно показал всю сложность поведения крысы в лабиринте (и других подобных ситуациях), особенно в начальном периоде решения задачи, когда решающую роль играет исследовательское поведение животного. Не менее ценным является в этой концепции то, что делается акцент на активность и инициативу животного и подчеркивается роль внутренних факторов (психического настроя животного) в процессе решения задач.
Опровергается тезис о «случайности» попыток решения задач и опытами с применением «латентного обучения». В таких опытах сравнивается скорость образования навыка у животных, которых непосредственно перед началом эксперимента помещают в лабиринт или «проблемный» ящик, с таковой у животных, которым предоставлялась возможность предварительно активно ознакомиться с установкой, побегать в ней. В последнем случае вырабатывается двигательный навык значительно быстрее.
В этой связи важно отметить, что каждая крыса, впервые попавшая в лабиринт, ведет себя иначе, особенно на первоначальной ориентировочной фазе, предшествующей решению задачи, когда пробежки совершаются еще без всякого подкрепления и служат лишь накоплению опыта. Варьирует даже модальность ведущей рецепции при первоначальном обследовании лабиринта (одни крысы руководствуются преимущественно визуальными, другие – кинестетическими стимулами и т.д.).
Крыса, просто бегая по лабиринту, познает его еще до решения задачи, связанного с получением пищевого подкрепления (или наказания) за правильную (или неправильную) ориентацию. Разумеется, индивидуальные особенности исследовательского поведения животного сказываются и в ходе самого решения задачи. Значительные индивидуальные различия поведения вообще являются характерным признаком процессов научения.
Итак, активная познавательная деятельность животного является важнейшей предпосылкой успешного формирования навыка при решении задач. В сущности, именно познавательный компонент определяет природу навыка. По Леонтьеву, важнейшим критерием навыка является выделение при решении задач особого состава или стороны деятельности, отвечающей условиям, в которых дан побуждающий деятельность животного предмет. Леонтьев обозначил этот компонент деятельности операцией. Выделение операции в двигательной активности животного и указывает на то, что мы действительно имеем дело с истинным навыком, а не с другой формой научения, поэтому Леонтьев считает навыками лишь закрепленные операции.
Выделение операции можно показать на простом опыте, проведенным А. В. Запорожцем и И. Г. Диманштейн по методу «обходного пути». В аквариум ставится поперечная перегородка из марли так, чтобы у боковой стенки оставался свободный проход. В меньшую часть аквариума в начале опыта помещается подопытная рыба, в большую, за перегородкой, – приманка (кусочки мяса). Чтобы заполучить приманку, подопытное животное должно обойти преграду (перегородку), что ему и удается после ряда безуспешных попыток найти дорогу к пище напрямик. В поисках пути к приманке животное производит локомоторные действия, в которых Леонтьев вычленяет двоякое содержание: 1) направленную деятельность, приводящую к результату и возникающую под влиянием свойства самого побуждающего деятельность предмета (запахи мяса), и 2) деятельность, связанную с воздействием преграды, т.е. с условиями, в которых дан побуждающий деятельность предмет. Именно эта деятельность и явится операцией.
После того как рыба (в описанном опыте – сомик) хорошо усвоила обходной путь, преграда была удалена, тем не менее рыба полностью повторяла прежний путь, как если бы преграда была на месте. Только постепенно путь рыбы к приманке выпрямлялся (рис. 15,а). Подобные опыты производились со сходными результатами и на крысах (рис. 15,6).
Следовательно, отмеченные компоненты деятельности здесь выступают еще слитно; воздействие, определяющее обходное движение, еще прочно связывается с воздействием пищи, с ее запахом. Перегородка не отделяется от приманки, воздействие преграды пока еще не воспринимается как свойство другой вещи – словом, операция здесь еще не выделяется.
По поводу описанного здесь опыта с сомиком Леонтьев пишет, что деятельность животных определяется уже фактически воздействием со стороны отдельных вещей (пища, преграда), но отражение действительности остается у них отражением совокупности отдельных ее свойств.
В приведенном примере познавательный компонент, а соответственно и навык в целом находятся на еще очень низком уровне; выученная животным траектория обходного пути оказалась настолько прочно закрепленной, что лишь постепенно элиминировалась в новых условиях среды (т.е. после удаления перегородки). Это типичный пример автоматизированного навыка. Чем сложнее навык, тем больше его познавательное значение и наоборот. В навыках высшего ранга, характерных для высших позвоночных, операция играет чрезвычайно важную познавательную роль. Однако и у этих животных закрепление индивидуального опыта совершается нередко в форме примитивных навыков наподобие описанного. Уровень навыка зависит в каждом конкретном случае от определяющих факторов биологии вида и от ситуации, в которой животное встречается с той или иной более или менее сложной задачей.
Рис. 15. Формирование обходного пути и его сохранение после удаления преград у сомика (а) и крысы (б). Пояснения см. в тексте
Говоря о структуре и динамике навыка, важно подчеркнуть, что понятие преграды не следует понимать лишь в физическом смысле, как перегородку на пути животного. Под преградой понимается любое препятствие к достижению побуждающего объекта («цели») при решении задачи. Это подчеркивал в свое время и Протопопов, который сумел экспериментально доказать, что любые двигательные навыки формируются у животных путем преодоления «преграды», что содержание навыков определяется именно характером самой преграды. Стимул же влияет, по Протопопову, на навык только динамически, определяя быстроту и прочность его закрепления.
Таким образом, преодоление преграды составляет наиболее существенный элемент формирования навыка не только при его. выработке описанным методом, но и при всех других методах, широко применяемых в зоопсихологических исследованиях. В частности, это относится к методам «лабиринта» и «проблемного ящика». Именно в способах преодоления преграды проявляется познавательная функция навыка.
Изучая познавательные аспекты формирования навыков у животных, венгерский зоопсихолог Л. Кардош также показал, что в ходе обучения в лабиринте у животного накапливается значительный запас информации. В результате уже в начале лабиринта животное «в памяти... видит дальше стен, закрывающих его поле ощущений; эти стены становятся как бы прозрачными. В памяти оно "видит" цель и наиболее важные с точки зрения локомоции (перемещения) части пути, открытые и закрытые двери, разветвления и прочее, "видит" точно так и там, где и как оно видело и в действительности во время обхода лабиринта».*
* Кардош Л. Некоторые проблемы сравнительно-психологического анализа процесса учения // Annales Universitatis Scientiarum Budapestinensis. Sect. Paedagogica et Psychologica. T.1. Budapest, 1970. P. 15.
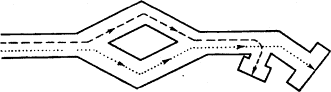
Рис. 16. Схема опыта Кардоша. Пояснения см. в тексте
Вместе с тем Кардош четко показал и границы познавательных возможностей животного в лабиринте, как и вообще при решении пространственно временных задач. Здесь имеются две возможности: локомоторное и манипуляционное познавание (второе происходит при формировании инструментальных навыков). В первом случае животное изменяет свое пространственное отношение к среде без того, чтобы сама среда изменилась. Если же в окружающей животное среде что-то изменяется в результате изменения поведения животного, то речь идет уже о манипуляторной активности животного. В исследованиях, произведенных вместе с И. Баркоци, Кардош показал, что с большим трудом можно крысу научить выбирать в одном и том же лабиринте разные пути, приводящие к одной точке, а затем двигаться дальше различным образом, например прямо или в сторону (рис. 16). Это пример локомоторного познавания. Но, согласно Кардошу, нельзя обучить животное (за исключением, возможно, человекообразных обезьян) тому, что в зависимости от выбора того или иного пути передвижения «случается то или нечто другое», т.е. произойдут изменения в окружающей среде (в экспериментах пища подменялась другим подкреплением – водой). Человек же, как пишет Кардош, «удивился бы, найдя в одном и том же месте разные предметы, когда он подходил справа и слева, но он обучился бы после первого же опыта. Развитие именно здесь делает скачок... Человек полностью может освободиться от направляющего воздействия пространственного порядка, если временно-причинные связи требуют другого».*
* Ibid. P. 19.
Научение и общение. Подражание
Существенную роль в формировании поведения высших животных играют явления подражания, которые в основном, хотя и не все, относятся к сфере научения. Мы здесь не будем рассматривать формы подражания, которые относятся к инстинктивному поведению. Это широко распространенная среди животных взаимная стимуляция – аллеломиметическое поведение (рис. 17), при котором выполнение видотипичных действий одними животными является побуждающим фактором для других. Последние в результате начинают выполнять такие же действия (одновременные отдых, собирание пищи и т.д.). Следовательно, в данном случае имеет место взаимопоощрение видотипичной деятельности.
Научение же путем подражания («имитационное научение») заключается в индивидуальном формировании новых форм поведения, но путем одного лишь непосредственного восприятия действий других животных. Таким образом, мы имеем здесь дело с научением на основе общения. Имитационное научение, как и всякое научение вообще, можно подразделить на облигатное и факультативное. При облигатном имитационном научении результат научения вполне укладывается в рамки видового стереотипа. Особенно это относится к молодым животным, которые путем подражания научаются выполнять некоторые жизненно необходимые действия обычного поведенческого «репертуара» своего вида. Так, у молоди стайных рыб защитная реакция на появление хищника (бегство) формируется в результате подражания поведению других рыб при одном лишь виде поедания хищником членов стаи. Л. А. Орбели считал такое имитационное поведение «главным охранителем вида», ибо «громадное преимущество заключается в том, что "зрители", присутствующие при акте повреждения члена их же стада или их сообщества, вырабатывают рефлекторные защитные акты и таким образом могут в будущем избежать опасности».* Облигатное имитационное научение является также важным элементом реакции следования (см. ч. II, гл. 3) и распознавания молодыми млекопитающими пищевых объектов. Путем облигатного имитационного научения молодые животные накапливают опыт в гнездостроении у птиц (В. П. Промптов) и шимпанзе (Дж. ван Лавик-Гудолл) и т.д.
* Орбели Л. А. Вопросы высшей нервной деятельности. М.; Л., 1949. С. 352.
Рис. 17. Схема категории подражания у животных (по Фабри)
Факультативное имитационное научение в простейших формах представлено в имитации невидотипичных движений на основе облигатного (аллеломиметического) стимулирования. Сюда относятся, например, случаи имитирования обезьянами действий человека, особенно при их содержании в домашней обстановке. Производимые ими при этом действия с предметами быта или инструментами, конечно, выходят за рамки видового поведения. Поскольку здесь имеет место научение новым приемам манипулирования, в данном случае можно говорить о невидотипичном имитационном манипулировании.
Высшим проявлением факультативного имитационного научения следует явно считать решение задач путем подражания (или хотя бы облегчение решения). При таком «имитационном решении задач» у животного-зрителя» вырабатывается определенный навык в результате одного лишь созерцания действий другой особи, направленных на решение соответствующей задачи. Способность к этому установлена у разных млекопитающих: человекообразных и низших обезьян, собак, кошек, крыс. У обезьян имитационное решение задач играет, очевидно, особенно большую роль. Советский исследователь поведения животных А. Д. Слоним, например, считает, что формирование условных рефлексов происходит в обезьяньем стаде преимущественно на основе подражания. В пользу этого говорят и полевые наблюдения, выполненные рядом исследователей в последние годы.
Правда, на основе имитационного научения у «зрителей», очевидно, не могут формироваться инструментальные навыки. Об этом свидетельствуют, в частности, опыты американского исследователя Б.Б.Бека, в которых павианы-«зрители» наблюдали за употреблением орудия при решении задачи у сородича. «Зрители» не оказались способными к имитационному решению столь сложной задачи, но впоследствии они намного чаще и интенсивнее, чем до опытов, манипулировали этим орудием. Это показывает активирующую роль аллеломиметического поведения и невидотипичного имитационного манипулирования при выработке сложных навыков в условиях общения.
Соотношения отдельных категорий и форм подражания у животных можно проиллюстрировать схемой (по К.Э. Фабри – см. рис. 17).
Эта схема неполная в том отношении, что не отражает явления подражания в области сигнализации и коммуникации (например, звукоподражание у птиц). Однако здесь нет принципиальных отличий от отмеченных общих закономерностей и характеристик, и поэтому данная классификация в соответствующих своих разделах вполне применима и для этой области. Так, например, к стимуляции видотипичного поведения -относится и стимуляция видотипичной акустической сигнализации («хоровое пение» у лягушек или птиц), невидотипичному имитационному манипулированию соответствует подражание чужим звукам и песням у птиц, а к облигатному имитационному научению относится, в частности, освоение птенцами видотипичных звуков путем подражания пению взрослых особей. Еще в 1773 г. английский ученый Д.Баррингтон воспитывал молодых коноплянок совместно с птенцами жаворонков. В результате коноплянки перенимали пение жаворонков. С тех пор аналогичные опыты ставились на птицах многократно.
Что же касается обезьян, то в опытах, поставленных Н. А. Тих на молодых гамадрилах, намечались, правда нечетко, некоторые намеки на возможность выработки даже таких навыков, которые можно было бы квалифицировать как имитационное решение задач на основе звукоподражания (в этих опытах обезьяны получали пищевое подкрепление за подражание звукам, которым в их присутствии обучали других обезьян).
Исходя из приведенной классификации можно определить два принципиально различных подхода к изучению подражания у животных.
Аллеломиметическое поведение изучается путем раздельного обучения изолированных друг от друга животных, которые затем сводятся вместе. Советский зоопсихолог Б. И. Хотин, например, обучал одних животных (рыб, птиц, млекопитающих) реагировать положительно на определенный сигнал, а других – отрицательно. После того как животные были сведены вместе и подвергнуты воздействию этого раздражителя, выяснилось, что преобладает: результаты обычного (неимитационного) научения или взаимная стимуляция. В результате можно судить о силе аллеломиметической реакции, т.е. инстинктивного подражания.
При изучении же имитационного научения животные с самого начала общаются между собой. Одна особь (или несколько особей) выполняет при этом роль «актера», т.е. обучается экспериментатором за соответствующее вознаграждение на виду у других особей – «зрителей». Если в результате последние без собственных упражнений и подкрепления вследствие одних лишь наблюдений за действием «актера» также научаются решать поставленную ему задачу, то можно говорить о факультативном имитационном научении и даже об имитационном решении задач. Так, например, в экспериментах с обезьянами все участвующие в опыте животные устремляются к месту его проведения и внимательно наблюдают, как вожак добывает и получает подкрепление. В итоге соответствующие навыки формируются у всех «зрителей», что проявляется в успешном решении ими той же задачи в отсутствие вожака.
На этом примере видно, как проявляется еще одна особенность, характерная для проявлений подражания в естественных условиях. Дело в том, что наряду с взаимным стимулированием к совместному выполнению определенных (инстинктивных) действий в сообществах животных действует и фактор противоположного рода – подавление действий членов сообщества «господствующими» особями. Так, в приведенном примере обезьяны боялись вплотную подойти к экспериментальной установке, а тем более брать пищу из кормушки. Вместе с тем у обезьян существуют особые «примиряющие» сигналы, оповещающие доминирующую особь о готовности «зрителей» «только смотреть», чем обеспечивается возможность осуществления аллеломиметического поведения и имитационного научения. Отсюда явствует, что явления подражания сложным образом переплетаются с внутригрупповыми отношениями животных.
* * *
Подводя итог обзору инстинктивного поведения и научения у животных, вернемся еще раз к описанной ранее общей схеме поведенческого акта. Эта схема, разумеется, не отражает всего многообразия вариантов поведенческих актов в их конкретных проявлениях.
Но во всех этих вариантах остается в силе основная закономерность: каждое действие животного начинается с внутреннего стимула, который в виде потребности активирует животное, кладет начало его поиску раздражителей. Это начало, как и общее- направление этого поиска, всегда генетически фиксировано, равно как и конец – заключительные движения животного в завершающей фазе как пройти этот путь от начала до конца между этими «жестко запрограммированными» основными элементами поведения – зависит от лабильных компонентов поведения животного, от его умения быстро и правильно ориентироваться во всех конкретных ситуациях, в которые оно попадает. Как мы видели, особое значение это имеет в начале пути.
У высших животных факультативное научение, сопряженное с формированием операций, является основным средством для продвижения к завершающей фазе. Но поскольку операция направлена на преодоление преграды, т.е. условий, в которых дан объект завершающего поведения, то здесь особенно важна более полноценная по возможности ориентировка животного во времени и пространстве. Ясно, что эта ориентировка будет тем более совершенной, чем выше уровень психической деятельности животного, его познавательные способности. Наибольший эффект обеспечивают в этом отношении высшие психические функции, особенно интеллектуальные способности, придающие всему поведению животного наибольшую гибкость и адаптивность. К этому вопросу мы еще вернемся при рассмотрении эволюции психики.
Часть II
РАЗВИТИЕ ПСИХИЧЕСКОЙ ДЕЯТЕЛЬНОСТИ ЖИВОТНЫХ В ОНТОГЕНЕЗЕ
Глава 1 ПРОБЛЕМА ОНТОГЕНЕЗА ПОВЕДЕНИЯ
Врожденное и приобретаемое в индивидуальном развитии поведения
Психическую деятельность можно познать лишь в процессе ее развития, и поэтому основное внимание зоопсихолога должно быть обращено на индивидуальное и историческое становление поведения животных. Именно так понимал изучение психической деятельности животных В.А.Вагнер, подчеркивая, что это изучение может быть строго научным лишь при применении двух методов, составляющих единство: онтогенетического, основанного на сравнении фактов из жизни особи, и филогенетического, основанного на сравнении фактов из жизни вида. Вместе эти два метода составляют единый биогенетический метод.
В этой части мы рассмотрим вопросы индивидуального, онтогенетического развития поведения; историческому, филогенетическому аспекту будет посвящена следующая часть.
Как было показано, при анализе любого проявления психической деятельности встает прежде всего вопрос о врожденных и приобретаемых компонентах поведения. Но, как показывает уже сама постановка вопроса, решение его по необходимости предполагает изучение онтогенеза поведения, которое только и позволяет судить о том, что же получает особь в наследство от предыдущих поколений в генетически фиксированном врожденном виде и чему она должна самостоятельно научиться в порядке приобретения индивидуального опыта. Таким образом, коренной вопрос зоопсихологии об инстинкте и научении во многих отношениях сливается с проблемой онотогенеза поведения.
Еще со времен Ламарка и Дарвина ученым было ясно, что поведение взрослого животного формируется из врожденных и благоприобретаемых элементов, но их удельный вес, взаимоотношения и изменения в ходе индивидуального развития служили предметом разногласий и горячих споров. Американский зоопсихолог Кллойд - Морган, например, дал в своей известной книге «Привычка и инстинкт», вышедшей в самом конце XIX в., глубокий анализ проблемы врожденного и приобретаемого в поведении животных, построенный целиком на онтогенетическом подходе. Он писал, что «деятельность, являющаяся результатом координирования подобранных (в результате индивидуального опыта. – К.Ф.) 10% первоначально бессвязных движений, есть новый продукт, и этот продукт есть результат усвоения, приобретения, а не наследуется в качестве определенного, координированного действия. Как скульптор создает статую из куска мрамора, так усвоение создает действие из массы данных случайных движений. Или как архитектор строит собор из неопределенной массы материала, выбирая надлежащие части, придавая им форму, соединяя их, так приобретение создает привычный облик из данного количества неопределенных движений, выбирая, видоизменяя эти движения и приводя их в известные отношения. Приобретается определенное, координированное, реактивное или ответное действие. Но, – продолжает Морган, – есть известные действия, которые определены с самого дня рождения, которые наследуются готовыми и сочетание или координирование которых тотчас после рождения уже отличается полным совершенством».* В качестве примера такого рода действий Морган ссылается на плавание молодой водяной курочки, впервые входящей в воду, или сооружение кокона шелковичным червем «без всякой предварительной практики или опыта». «Определенность и координирование действий, – пишет он, – в данном случае не индивидуальны, а заимствованы от предков».*
* Ллойд-Морган К. Привычка и инстинкт. Спб., 1899. С. 21–22.
** Там же. С. 22.
В отличие от такого всестороннего подхода к проблеме онтогенеза поведения животных, при котором учитываются как генетически фиксированные, так и индивидуально приобретаемые его компоненты и подчеркивается значение деятельности как ведущего фактора отбора и модификации врожденных элементов поведения наряду с созреванием действий, не нуждающихся в научении, позднее, особенно в 20–30-е годы нашего века, неоднократно делались попытки одностороннего решения вопроса, сведения всего процесса развития поведения к действию лишь одной из этих категорий факторов. Так, Г.Е.Когхилл и Цин Янг Куо в Америке, В.М.Боровский в Советском Союзе отстаивали точку зрения, что все в поведении является благоприобретенным, что животное должно всему научиться, причем начиная уже с эмбрионального периода. Этот взгляд, в модифицированном виде подчас встречающийся и поныне, являлся реакцией на переоценку генетических факторов и недооценку факторов среды в онтогенезе животных. Куо, к работам которого мы еще вернемся, считал, например, возможным писать о «психологии без наследственности». Боровский при изложении своей концепции исходил из необходимости борьбы против телеологических взглядов на развитие организмов, согласно которым все в будущей жизни организма заранее предопределено изначально действующими внутренними факторами.
Корни механистического взгляда, всецело отрицающего наличие и роль внутренних факторов в развитии организма, уходят к философии Декарта, пытавшегося приложить принцип действия машины к поведению животных. Но в конце концов такое объяснение отличается от телеологического лишь тем, что изначально действующие движущие силы выносятся из организма вовне.
Современное понимание взаимоотношений врожденного и благоприобретенного в онтогенезе поведения животных исходит из признания не только наличия, но и взаимообусловленности этих компонентов. В противоположность вульгарно-материалистическому толкованию процесс онтогенеза поведения открывается нам во всей его сложности и противоречивости как подлинно диалектическое формирование качественно нового в результате количественных преобразований первичных функциональных состояний развивающегося организма. Такое диалектико-материалистическое понимание онтогенеза характеризует, в частности, теорию системогенеза П.К. Анохина.
Взаимопроникновение и различные сочетания безусловно- и условнорефлекторных элементов в онтогонезе поведения побудили Л.В.Крушинского выдвинуть тезис об «унитарных реакциях», под которыми он понимает акты поведения, имеющие сходное внешнее выражение при различных способах их формирования. Унитарные реакции, по Крушинскому, представляют собой «единые, целостные акты поведения, в которых объединены, интегрированы условные и безусловные рефлексы»,* они направлены «к выполнению определенного акта поведения, имеющего разные пути осуществления и в то же время определенный шаблон конечного исполнения».** Соотношение условных и безусловных рефлексов в унитарной реакции не строго фиксировано, а сама она направлена на выполнение одиночного приспособительного действия. В ходе онтогенеза унитарные реакции интегрируются, по Крушинскому, в форме «многоактного поведения», связанного с обеспечением основных биологических потребностей организма. Эти формы поведения не являются, однако, простой суммой унитарных реакций, а обладают гибкой структурой, что позволяет животному приспосабливаться в процессе своего развития к самым различным условиям жизни.
* Крушинский Л.В. формирование поведения животных в норме и патологии. М., 1960. С. 10.
** Там же. С. 11.
Биологическая обусловленность онтогенеза поведения животных
Процессы, совершающиеся в ходе онтогенеза, во многом обнаруживают те же закономерности, что и процессы филогенеза. Для понимания сущности формирования индивидуального поведения первостепенное значение имеет тот факт, что морфофункциональные преобразования, характеризующие эволюцию животного мира, играют не меньшую роль и в онтогенезе. Так, известный советский зоолог Б.С. Матвеев показал, что на разных этапах онтогенеза позвоночных имеют место явления смены функций, о чем еще пойдет речь. Им же были глубоко проанализированы взаимоотношения развивающегося организма с окружающей средой и выявлены важные закономерности, вытекающие из этого взаимодействия. Изучив преобразования организации животных на разных стадиях индивидуального развития, Матвеев пришел к выводу, что отношения организма к условиям среды претерпевают в ходе онтогенеза существенные изменения и на разных стадиях развития организмы приспосабливаются к среде по-разному. Особенно на ранних стадиях онтогенеза возможна далеко идущая адаптивная перестройка в морфофункциональной сфере. Ведущими, по Матвееву, являются функциональные изменения экзосоматических, т.е. внешних, «рабочих», органов, которые приводят к корреляционным сдвигам во всем организме как единой целостной системе.
Эти выводы приобретают особое значение для познания развития психической деятельности, если вспомнить, что врожденные двигательные координации, инстинктивные движения являются функцией экзосоматических органов и обусловливаются морфологическими признаками.
Важным моментом, определяющим ход онтогенеза, является степень зрелорождения животных. Способность к самостоятельному выполнению жизненных функций в весьма различной мере выражена у разных новорожденных, что обусловлено различиями в филогенетическом уровне и образе жизни. Отсюда проистекают существенные различия в онтогенезе поведения.
В еще большей мере разнообразие онтогенеза у разных групп животных определяется наличием или отсутствием личиночных форм, ведущих подчас совершенно иной образ жизни, чем половозрелые формы (имаго). Для формирования психического отражения в этом отношении весьма существенно, что личиночные и имагинальные формы зачастую по-разному передвигаются и вообще обладают в корне различными двигательными способностями. Достаточно вспомнить соответствующие различия между лягушкой и головастиком. В процессе метаморфоза, превращающего личинку в половозрелую особь, происходят сложнейшие преобразования поведения. Чрезвычайно велики различия онтогенеза между позвоночными и беспозвоночными животными. У последних личиночные стадии являются общераспространенными и в большинстве случаев поведение личинки и имаго имеет мало или даже не имеет ничего общего.
* * *
Сложность и многообразие проявлений онтогенеза поведения животных чрезвычайно затрудняет периодизацию этого процесса. Схемы периодизации, предложенные разными авторами, являются слишком дробными и применимы поэтому только для определенных групп животных (нельзя, например, по наблюдениям над развитием кролика или собаки судить о стадиях онтогенеза поведения у животных «вообще»).
По этой причине мы воздержимся при дальнейшем изложении от детальной периодизации развития психической деятельности. Наиболее целесообразным и отражающим решающие вехи в онтогенезе поведения мы считаем выделение лишь трех крупных периодов: пренатального, раннего постнатального и ювенильного (игрового).
Последний период, предшествующий половому созреванию, встречается не у всех животных, а только у тех, которым свойственна игровая активность. Как будет дальше показано, эта активность играет исключительно большую роль в онтогенезе высших животных. Что же касается игры как критерия психического развития, то сошлемся на выдающегося исследователя поведения насекомых, лауреата Нобелевской премии, австрийского ученого К.Фриша, который подчеркивает, что отсутствие игры у насекомых и ее наличие у высших позвоночных указывают на знаменательное различие между этими группами животных и свидетельствуют о перевесе наследуемых форм поведения у первых и индивидуального приобретения опыта у вторых.
Аналогичным образом, очевидно, игра выступает как веха и в онтогенезе поведения, более обстоятельное ознакомление с которым мы, однако, начнем с изначального, эмбрионального периода.
Глава 2 РАЗВИТИЕ ПСИХИЧЕСКОЙ ДЕЯТЕЛЬНОСТИ В ПРЕНАТАЛЬНОМ ПЕРИОДЕ
Врожденное и приобретаемое в пренатальном развитии поведения
Морфофункциональные основы эмбриогенеза поведения
Поведение эмбриона является во многих отношениях основой всего процесса развития поведения в онтогенезе. Как у беспозвоночных, так и у позвоночных установлено, что развивающийся организм производит еще в пренатальном (дородовом) периоде движения, которые являются элементами будущих двигательных актов, но еще лишены соответствующего функционального значения, т.е. еще не могут играть приспособительную роль во взаимодействии животного со средой его обитания. Эта функция появляется лишь в постнатальном периоде его жизни. В таком смысле можно говорить о преадаптационном значении эмбрионального поведения.
Академик И.И.Шмальгаузен выделил несколько типов онтогенетических корреляций, среди которых особый интерес для познания закономерностей развития поведения представляют «эргонтические корреляции» (или функциональные в узком смысле), т.е. соотношения между частями и органами, обусловленные функциональными зависимостями между ними. Шмальгаузен при этом имел в виду типичные («окончательные») функции органов. Таковыми являются уже на ранних стадиях эмбриогенеза, например, функции сердца или почек зародыша. В качестве характерного примера эргонтических корреляций Шмальгаузен указывает и на зависимость между развитием нервных центров и нервов и развитием периферических органов – органов чувств и конечностей. При экспериментальном удалении этих органов соответствующие элементы нервной системы недоразвиваются.
Как установил еще в 30-е годы Б.С.Матвеев, эмбриогенез характеризуется коррелятивными сдвигами в соотношениях между развивающимися органами, которые являются следствием нарушения взаимосвязей частей зародыша в развивающемся организме. В результате возникают функциональные изменения, определяющие характер деятельности формирующихся органов.
Эти выводы ведущих советских зоологов показывают всю сложность морфофункциональных связей и взаимозависимостей, определяющих формирование поведения животных в эмбриогенезе. Это нашло свое отражение и в концепции системогенеза П.К. Анохина. «Развитие функции, – говорил Анохин, – идет всегда избирательно, фрагментарно в отдельных органах, но всегда в крайней согласованности одного фрагмента с другим и всегда по принципу конечного создания работающей системы».* В частности, на примерах развития эмбрионов кошки, обезьяны и человеческого плода он показал функциональную обусловленность асинхронности развития отдельных структур зародышей, подчеркнув, что «в процессе эмбриогенеза идет ускоренное созревание отдельных нервных волокон, которые определяют жизненные функции новорожденного» (например, сосательные движения), ибо для его выживания «"система отношений" должна быть полноценной к моменту рождения».**
* Анохин П.К. О морфологических закономерностях развития функции в эмбриогенезе животных и человека // Труды 6-го Всесоюзного съезда анатомов и эмбриологов. Т. 1. Харьков, 1961. С. 25.
** Там же. С. 27–28.
Как показали исследования А.Д.Слонима и его сотрудников, внутриутробные движения влияют на координацию физиологических процессов, связанных с мышечной деятельностью, и тем самым способствуют подготовке поведения новорожденного. По данным Слонима, новорожденные козлята и ягнята способны, не утомляясь, бегать до двух часов подряд. Эта возможность обусловлена тем, что в ходе эмбриогенеза путем упражнения сформировалась координация всех функций, в том числе и вегетативных, необходимых для осуществления такой интенсивной деятельности уже в самом начале постнатального развития. В частности, к моменту рождения уже скоординирована регуляция минутного объема сердца и частота дыхания, как и других физиологических функций.
Говоря о принципе «конечного создания работающей системы» (Анохин) или эмбриональной преадаптации постнатального поведения, т.е. о преадаптационном значении эмбрионального поведения, нельзя забывать об относительности этих понятий. Эмбриональную преадаптацию нельзя понимать как некую изначальную, «фатальную» предопределенность. Условия жизни взрослого животного, его взаимодействие с компонентами окружающей среды в конечном итоге определяют условия развития зародыша, функционирование его развивающихся органов. Общие же закономерности и направление развития функции детерминированы сложившимися в филогенезе и генетически фиксированными факторами.
Эмбриональное научение и созревание
В указанной связи приобретает особый вопрос об «эмбриональном научении», считавшемся некоторыми исследователями преимущественным, если не единственным фактором всего сложного процесса первоначального формирования экзосоматических функций. К числу этих исследователей относится уже упоминавшийся известный американский ученый Цин Янг Куо, который еще в 20–30-х годах нашего века пытался объяснить весь процесс формирования поведения животных исключительно накоплением двигательного опыта и изменениями в окружающей зародыш среде. Куо сумел одним из первых убедительно показать, что уже в ходе эмбриогенеза происходит упражнение зачатков будущих органов, постепенное развитие и совершенствование двигательных функций путем накопления «эмбрионального опыта».
Куо изучал эмбриональное поведение на многих сотнях куриных зародышей (рис. 18). Чтобы получить возможность непосредственно наблюдать за движениями эмбрионов, он разработал и осуществил поистине виртуозные операции: перемещал зародыш внутри яйца, вставлял окошки в скорлупу и т.д. Ученый установил, что первые движения зародыша цыпленка, наблюдающиеся уже на четвертый, а иногда и на третий день инкубации, – это движения головы к груди и от нее. Уже через сутки голова начинает поворачиваться в стороны, и эти новые движения головы вытесняют прежние к 6–9-м суткам.
Рис. 18. Развитие куриного эмбриона. Сверху вниз: первые сутки инкубации (стадия первичной полоски – формируются зачатки головного и спинного мозга, скелета и скелетной мускулатуры), третьи сутки (зародыш на сосудистом поле желточного мешка), девятые сутки, пятнадцатые сутки
|
|
Очевидно, причина тому – отставание роста шейной мускулатуры от роста головы, которая к десятому дню составляет уже более 50% веса всего зародыша. Такую огромную голову мышцы могут только поворачивать, но не поднимать и опускать. Кроме того, по Куо, движения головы формируются под влиянием таких моментов, как ее положение по отношению к скорлупе, расположение желточного мешка, сердцебиение и даже движения пальцев ног, поскольку последние во второй половине инкубационного периода располагаются слева и сзади головы.
Во всем этом Куо видел проявления действия «анатомического фактора» развития поведения. Сходным образом специфические условия морфоэмбриогенеза, анатомические изменения, связанные с ростом и развитием зародыша, влияют, по его мнению, и на формирование других движений куриного зародыша. В итоге вылупившийся из яйца цыпленок обладает целым набором выработанных в период эмбриогенеза реакций, «механизмов», но они не направлены на необходимые для поддержания жизни раздражители. Широкая генерализация характерна, по Куо, как для раздражителей (нет избирательного отношения к ним, на весьма различные раздражители следует одинаковая реакция), так и для двигательных реакций (всегда двигается все тело, движения отдельных его частей еще плохо или совсем не координированы, они нецелесообразны, неэкономичны).
Из своих исследований Куо вывел заключение, что цыпленок должен всему научиться, что ни одна его реакция не появляется в готовом виде, а следовательно, нет врожденного поведения. Сорок лет спустя после опубликования своих первых работ Куо, уточняя свою точку зрения, указал на то, что генетически фиксированные предпосылки формирования поведения могут по-разному реализоваться в зависимости от конкретных условий развития зародыша, но важнейшую роль играет в этом процессе «расшифровки» генетической информации отношение эмбриона к окружающей его среде. Одновременно Куо подчеркивает, что эмбриональное научение не следует рассматривать в традиционном аспекте, поскольку в развитии поведения эмбриона существенную роль играет самостимулирование. Однако, как показывают результаты современных исследований, тактильная и проприоцептивная стимуляция, как представлял себе ее Куо, играет в развитии моторики в нормальном эмбриогенезе, очевидно, подчиненную роль.
Односторонний подход к проблеме становления поведения в онтогенезе, игнорирование врожденной основы индивидуального поведения, в том числе на эмбриональной стадии развития, являются, конечно, глубоко ошибочными. Если и можно говорить о некоем научении в период эмбриогенеза, то оно не происходит на пустом месте, а является развитием и видоизменением определенного генетического зачатка, воплощением и реализацией в индивидуальной жизни особи видового опыта, накопленного в процессе эволюции. Филогенез подготовил возможность развития поведения в онтогенезе в биологически полезном для особи и вида направлении. Наследственность проявляется не только в строении организма, его систем и органов, но и в их функциях.
Наследственная основа эмбриогенеза поведения выступает особенно отчетливо в тех случаях, когда элементы поведения новорожденного проявляются сразу как бы в «готовом виде», хотя возможность предшествующего «эмбрионального научения» исключается. К таким случаям, очевидно, относятся у млекопитающих поиск соска и сосущие движения новорожденных, звуковые реакции и др. Здесь можно говорить лишь о пренатальном созревании функции без эмбрионального упражнения, т.е. без пренатальной функциональной тренировки соответствующих морфологических структур. Для такого созревания явно достаточно одной лишь врожденной программы развития, возникшей и закрепившейся в ходе эволюции вида.
Хорошим примером, показывающим наличие и роль генетически фиксированных «программ действия», может служить поведение новорожденного кенгуру, который появляется на свет на столь незрелой стадии развития, что его можно сравнить с эмбрионом высших млекопитающих. В известной степени можно считать, что окончательное развитие эмбриона завершается в сумке матери. Но несмотря на состояние крайнего незрелорождения, детеныш совершенно самостоятельно перебирается в сумку матери, проявляя при этом поразительные двигательные и ориентационные способности. При этом нахождение сумки осуществляется на основе отрицательного гидротаксиса (а возможно, и хемотаксиса): оказавшись вне родовых путей, новорожденный, цепляясь за шерсть, поднимается по сухим ее участкам к сумке, находит ее вход, вползает в нее, находит там сосок, крепко присасывается к нему и остается длительное время висеть на нем. Шерсть матери смачивается перед этим на прихвостовых участках туловища родовыми водами, выливающимися после разрыва зародышевых оболочек, «дорожка» же, ведущая к сумке, остается сухой.
Здесь обращает на себя внимание строгая последовательность врожденных реакций. На эмбриональном этапе своего развития, который здесь резко укорочен, если не сказать прерван, детеныш не мог научиться ни отдельным поведенческим актам этой цепи, ни этой последовательности. До разрыва родовых оболочек он постоянно находился во влажной среде и не мог, таким образом, упражнять отрицательное гидротаксисное поведение. Никогда он не соприкасался с сухими объектами, в том числе с шерстью. Аналогично обстоит дело и с другими компонентами этого сложного поведенческого комплекса, формирование которых также невозможно объяснить «пренатальным научением».
Существенным для понимания процессов созревания элементов поведения в эмбриогенезе является то обстоятельство, что у позвоночных иннервация соматической мускулатуры туловища и конечностей предшествует замыканию рефлекторных дуг (у куриного эмбриона это замыкание происходит уже на 6–7-й день с начала инкубации). Однако сокращения этих мышц начинаются уже с момента их иннервации и носят, таким образом, на первых порах нерефлекторный характер. Эти движения являются ритмичными, так как они обусловливаются спонтанными нейрогенными ритмами («импульсными ритмами»). Эндогенная ритмика в нервно-мышечных структурах сохраняется на протяжении всей жизни животного и представляет собой один из важных факторов созревания элементов врожденного поведения.
Все же едва ли можно говорить о некоем «чистом» созревании поведенческих актов, особенно если иметь в виду упомянутые выше коррелятивные морфофункциональные связи. К тому же, вероятно, никогда не удастся полностью исключить возможность и непосредственного пренатального упражнения тех или иных двигательных элементов поведенческого акта. Поэтому, возражая против одностороннего постулирования «эмбрионального научения» как единственного фактора пренатального развития поведения, невозможно соглашаться и с противоположной крайней точкой зрения, однобоко акцентирующей внимание на процессах эмбрионального (и постнатального) созревания. Эту точку зрения сформулировал видный американский исследователь онтогенеза поведения животных Л.Кармайкл, который допускал, что даже у человека поведение является на «девять десятых» врожденным. В том и другом случае неверным является противопоставление врожденного и приобретаемого в поведении, о чем уже многократно говорилось.
Что же касается термина «эмбриональное научение», то, очевидно, более точным будет выражение «эмбриональная тренировка», во всяком случае, когда речь идет о ранних стадиях эмбриогенеза. На первых стадиях, как уже упоминалось, даже еще нет рефлекторных дуг; сомнительно, чтобы условнорефлекторные связи образовались и на средних стадиях эмбриогенеза. Очевидно, отсутствуют на этих этапах и явления привыкания. Однако в каждом случае функционирование развивающегося органа или системы, безусловно, является индивидуальным «усвоением» и «прилаживанием» видового опыта, т.е. инстинктивного поведения, в виде тренировки. Последняя, разумеется, также относится к категории научения как одна из его элементарных форм. Полноценное же научение, как еще будет показано, встречается лишь на завершающих этапах эмбриогенеза.
Сравнительный обзор развития двигательной активности зародышей
Беспозвоночные
Эмбриональное поведение беспозвоночных еще очень слабо изучено. Те немногие сведения, которые пока получены, относятся в основном к кольчатым червям, моллюскам и членистоногим. Известно, например, что зародыши головоногих моллюсков уже на ранних стадиях своего развития вращаются внутри яйца вокруг своей оси со скоростью один оборот в час. В других случаях зародыши передвигаются от одного полюса яйца к другому. Все эти движения осуществляются с помощью ресничек, что представляет особый интерес, если учесть, что этот примитивный способ передвижения у взрослых головоногих моллюсков отсутствует, но широко распространен среди личинок морских беспозвоночных.
Заслуживает также внимания, что к концу эмбриогенеза у беспозвоночных оказываются уже вполне сформированными некоторые инстинктивные реакции, имеющие первостепенное значение для выживания. У мизид (морские ракообразные), например, к моменту вылупления уже вполне развита реакция избегания, т.е. уклонения от неблагоприятных воздействий. Но, как показал канадский исследователь М.Беррилл, первоначально у эмбриона наблюдается спонтанное «вздрагивание» и «подергивание», которые затем все чаще возникают в ответ на прикосновение к икринке. Таким образом, первоначально спонтанные движения постепенно сменяются рефлекторными.
У других ракообразных – морских козочек (отряд бокоплавы) – с 11-го по 14-й день развития, т.е. до вылупления, наблюдаются спонтанные и ритмичные движения головы и других частей эмбриона, из которых впоследствии формируются специфические двигательные реакции этих рачков. Только к концу эмбриогенеза, в день вылупления, появляются двигательные ответы на тактильные раздражения (прикосновения волоском к зародышу, с которого в эксперименте была снята яйцевая оболочка). В естественных условиях весь набор движений взрослой особи обнаруживается уже спустя 10 часов после вылупления.
У эмбриона дафнии антенны, служащие у взрослых особей для плавания, начинают двигаться на средних этапах эмбриогенеза, а перед его завершением они поднимаются и принимают то положение, которое необходимо для выполнения плавательных движений. В это время (за несколько часов до выхода из яйца) антенны начинают особенно интенсивно двигаться.
В этих примерах отчетливо проявляется постепенное формирование рефлекторного ответа на основе первоначально эндогенно обусловленных движений, которые впоследствии связываются с внешними раздражителями, отчасти уже путем «эмбрионального научения». Этот процесс сопряжен с глубокими морфологическими преобразованиями.
Низшие позвоночные
Первые движения зародышей рыб, по данным ряда исследователей, также возникают спонтанно на эндогенной основе. Еще в 20-х годах было показано, что движения зачатков органов появляются в строгой последовательности, в зависимости от созревания соответствующих нервных связей. После появления сенсорных нервных элементов на поведение зародыша начинают влиять и экзогенные факторы (например, прикосновения), которые сочетаются с генетически предопределенной координацией движений. Постепенно первоначально генерализованные движения зародыша дифференцируются.
Вообще у зародышей костистых рыб обнаруживаются к концу эмбриогенеза такие движения: дрожание, подергивание отдельных частей тела, вращение, змеевидное изгибание. Кроме того, перед вылуплением производятся своеобразные «клевательные» движения, облегчающие выход из яйцевидной оболочки. Кроме того, выклеву способствуют и изгибательные движения тела. В ряде случаев удалось установить четкую связь между появлением новых двигательных актов и общим анатомическим развитием.
Сходным образом совершается формирование эмбрионального поведения и у земноводных. Из первоначально генерализованного сгибания всего тела зародыша постепенно формируются плавательные движения, движения конечностей и т.д., причем и здесь двигательная активность развивается первично на эндогенной основе.
Интересный пример представляет в этом отношении жаба Eleutherodactylus martinicensis с острова Ямайка, у которой выход из икринки как бы задерживается и личинка развивается внутри яйцевых оболочек. Тем не менее у нее проявляются все движения, свойственные свободноплавающим личинкам (головастикам) других бесхвостых земноводных. Как и у последних, плавательные движения формируются у этой личинки постепенно из более генерализованных двигательных компонентов: первые движения конечностей еще слиты с общим извиванием всего тела, но уже спустя сутки можно вызвать одиночные рефлекторные движения одних конечностей независимо от движений мышц туловища; несколько позже и в строгой последовательности появляются более дифференцированные и согласованные движения всех четырех конечностей, и наконец возникают во всех деталях вполне координированные плавательные движения с участием всех соответствующих моторных компонентов, хотя плавать сформировавшаяся к этому времени личинка еще не начинала, ибо она по-прежнему заключена в яйцевые оболочки.
Что же касается хвостатых амфибий, то Когхилл показал в свое время, что эмбрион амбистомы производит плавательные движения еще задолго до вылупления, изгибаясь сперва наподобие буквы «С» и позже как буква «S». Еще позже появляются движения ног, типичные для передвижения по суше взрослой амбистомы, причем нейромышечная система, детерминирующая первичные плавательные движения, определяет и эти локомоторные элементы, особенно последовательность и ритм движений. Кармайкл сумел на этом же объекте доказать, что этот механизм созревает без научения. Он вырастил эмбриона амбистомы в анестезирующем растворе ацетонхлороформа, который полностью обездвиживал зародыш, но не препятствовал его росту и морфогенезу. В таких условиях локомоторные способности развивались вполне нормально, и в итоге они не отличались от таковых контрольных организмов, развитие которых совершалось в обычных условиях. Из своих опытов Кармайкл вывел заключение, что формирование способности к плаванию не нуждается в процессах научения, а зависит исключительно от анатомического развития.
Однако, как справедливо отмечал по поводу этих опытов польский зоопсихолог Я. Дембовский, у подопытных эмбрионов подавлялись лишь мышечные движения, возможность накопления двигательного эмбрионального опыта, но не другие функции, в частности процессы в развивающейся нервной системе. Невозможно, как пишет Дембовский, создать такие экспериментальные условия, при которых нервная система развивалась бы, не функционируя. Поскольку нервная система начинает функционировать еще до того, как она окончательно сформировалась, это функционирование также является своего рода упражнением, которое в свою очередь является важным фактором развития нервной деятельности, а тем самым всего поведения зародыша.
Для выявления эндогенной обусловленности формирования двигательной активности зародышей производились интересные опыты на эмбрионах саламандр: пересаживались зачатки конечностей таким образом, что последние оказывались повернутыми в обратную сторону. Если бы формирование их движений определялось эмбриональным упражнением (путем афферентной обратной связи), то в ходе эмбриогенеза должна была бы произойти соответствующая функциональная коррекция, восстанавливающая способности к нормальному поступательному движению. Однако этого не произошло, и после вылупления животные с повернутыми назад конечностями пятились от раздражителей, которые у нормальных особей обусловливают движение вперед. Сходные результаты были получены и у эмбрионов лягушек: перевертывание зачатков глазных яблок на 180° привело к тому, что оптокинетические реакции оказались у этих животных смещенными в обратном направлении (рис. 19).
|
|
|
|
Рис. 19. Инверсии оптокинетической реакции у лягушек (опыт Сперри). Большие черные стрелки обозначают направление оптического раздражения, вызывающего оптокинетическую реакцию (маленькая светлая стрелка) в норме (вверху) и после поворота зачатков глазных яблок на 1800 (внизу)
Все эти данные приводят к заключению, что формирование в эмбриогенезе локомоторных движений и оптомоторных реакций (а также некоторых других проявлений двигательной активности) происходит у низших позвоночных, очевидно, не под решающим влиянием экзогенных факторов, а в результате эндогенно обусловленного созревания внутренних функциональных структур.
Птицы
Эмбриональное поведение птиц изучалось преимущественно на зародышах домашней курицы. Уже в конце вторых суток появляется сердцебиение, а начало спонтанной двигательной активности куриного эмбриона приурочено к 4-му дню инкубации. Весь период инкубации длится три недели. Движения начинаются с головного конца и постепенно простираются к заднему, охватывая все большие участки тела зародыша. Самостоятельные движения органов (конечностей, хвоста, головы, клюва, глазных яблок) появляются позже.
Как уже говорилось, Куо установил наличие и показал значение эмбриональной тренировки у зародыша курицы (а также других птиц), но впал при этом в односторонность, отрицая наличие врожденных компонентов поведения и спонтанной активности как таковой. В дополнение к сказанному приведем еще несколько примеров из работ этого выдающегося исследователя.
Куо установил, что максимальная двигательная активность эмбриона совпадает по времени с движениями амниона, т.е. внутренней зародышевой оболочки, обволакивающей зародыш. Из этого Куо заключил, что пульсация оболочки обусловливает начато двигательной активности последнего. Впоследствии, однако, В. Гамбургером было показано, что нет подлинной синхронизации между этими двигательными ритмами, а Р. В. Оппенгейм экспериментально доказал, что движения эмбриона не только не зависят от движения амниона, а, возможно, даже сами обусловливают их.
Формирование клевательных движений, по Куо, первично определяется ритмом биения сердца зародыша, ибо первые движения клюва, его открывание и закрывание, совершаются синхронно с сокращениями сердца. Впоследствии эти движения коррелируются со сгибательными движениями шеи, а незадолго до вылупления клевательный акт следует за любым раздражением тела в любой его части. Таким образом, реакция клевания, сформировавшаяся путем эмбриональной тренировки, имеет к моменту вылупления птенца весьма генерализованный характер. «Сужение» реакции в ответ на действие лишь биологически адекватных раздражителей происходит на первых этапах постэмбрионального развития. По Куо, так обстоит дело и с другими реакциями.
Важную роль в формировании эмбрионального поведения Куо отводил также непосредственному влиянию среды, окружающей зародыш, и происходящим в ней изменениям. Так, например, у куриного зародыша начиная с 11-го дня инкубации желточный мешок начинает надвигаться на брюшную сторону зародыша, все больше стесняя движения ног, которые до вылупления сохраняют согнутое положение, причем одна нога располагается поверх другой. По мере того как желток всасывается зародышем, верхняя нога получает все большую свободу движения, однако находящаяся под ней нога по-прежнему лишена такой возможности. Лишь после того как верхняя нога получит возможность достаточно далеко отодвигаться, нижняя может начать двигаться.
Таким образом, движения ног развиваются с самого начала не одновременно, а последовательно. Куо считал, что именно этим определяется развитие механизма последовательных движений ног цыпленка при ходьбе, что здесь кроется первопричина того, что вылупившийся из яйца цыпленок передвигается шагами, попеременно переставляя ноги, а не прыжками, отталкиваясь одновременно обеими лапками. Поддерживая концепцию Куо, Боровский писал по этому поводу, что «цыпленок шагает не потому, что у него имеется "инстинкт шагания", а потому, что иначе не могут работать его ноги и их механизм, выросшие и развившиеся в таких именно условиях. Механизм этот не появится как нечто готовое в тот момент, когда он понадобится, а развивается определенным путем, под влиянием других факторов».*
* Боровский В.М. Проблема инстинкта. Крымское гос. изд-во, 1941. С.133.
Многие механистические концепции Куо не выдержали экспериментальной проверки, произведенной более поздними исследователями, другие еще нуждаются в такой проверке, тем более что вскрытые им коррелятивные отношения не обязательно указывают на причинные связи. Не подтвердилось, в частности, мнение Куо о том, что ведущим фактором двигательной активности зародыша в раннем эмбриогенезе является сердцебиение.
Гамбургером и его сотрудниками было установлено, что уже на ранних стадиях эмбриогенеза движения зародыша имеют нейрогенное происхождение. Электрофизиологические исследования показали, что уже первые движения обусловливаются спонтанными эндогенными процессами в нервных структурах куриного эмбриона. Спустя 3,5–4 дня после появления первых его движений наблюдались первые экстероцептивные рефлексы, однако, Гамбургер, Оппенгейм и другие показали, что тактильная, точнее, тактильнопроприоцептивная стимуляция не оказывает существенного влияния на частоту и периодичность движений, производимых куриным эмбрионом на протяжении первых 2–2,5 недель инкубации. По Гамбургеру, двигательная активность зародыша на начальных этапах эмбриогенеза «самогенерируется» в центральной нервной системе.
Гамбургером производился следующий опыт: перерезав зачаток спинного мозга в первый же день развития куриного эмбриона, он регистрировал впоследствии (на 7-й день эмбриогенеза) ритмичные движения зачатков передних и задних конечностей. Нормально эти движения протекают синхронно. У оперированных же эмбрионов эта согласованность нарушилась, но сохранилась самостоятельная ритмичность движений.
Это указывает на независимое эндогенное происхождение этих движений, а тем самым и соответствующих нервных импульсов, на автономную активность процессов в отдельных участках спинного мозга. С развитием головного мозга он начинает контролировать эти ритмы. Вместе с тем эти данные свидетельствуют о том, что двигательная активность не обусловливается исключительно обменом веществ, например, такими факторами, как уровни накопления продуктов обмена веществ или снабжения тканей кислородом, как это принималось некоторыми учеными.
При изучении эмбрионального развития поведения птиц необходимо учитывать специфические особенности биологии исследуемого вида, которые отражаются и на протекании эмбриогенеза. Особенно это касается различий между выводковыми (зрелорождающимися) и птенцовыми (незрелорождающимися) птицами. Так, например, как показал советский исследователь Д. Н. Гофман, по сравнению с курицей грач развивается более ускоренно, быстрее накапливается масса тела зародыша, зато у курицы эмбриогенез проходит более равномерно и имеется больше периодов роста и дифференциации. Последний период формирования морфологических структур и поведения проходит у курицы еще внутри яйца, у грача же (как не зрелорождающейся птицы) этот период относится к постэмбриональному развитию.
Млекопитающие
В отличие от рассмотренных до сих пор животных зародыши млекопитающих развиваются в утробе матери, что существенно усложняет (и без того весьма трудное) изучение их поведения, поэтому по эмбриональному поведению млекопитающих накоплено значительно меньше данных, чем по куриному эмбриону и зародышам земноводных и рыб. Непосредственные визуальные наблюдения возможны лишь на эмбрионах, извлеченных из материнского организма, что резко искажает нормальные условия их жизни. Рентгенологические исследования указывают на то, что двигательная активность таких искусственно изолированных зародышей выше, чем в норме. Вместе с тем именно на таких объектах, преимущественно зародышах грызунов, были получены те данные, которыми мы сегодня располагаем.
Так, например, по Кармайклу, развитие двигательной активности совершается у зародыша морской свинки следующим образом. Первые движения состоят в подергивании шейно-плечевого участка туловища эмбриона. Они появляются приблизительно на 28-й день после оплодотворения. Постепенно появляются и другие весьма разнообразные движения, а к 53-му дню, т.е. приблизительно за неделю до родов, формируются четко выраженные реакции, которые достигают максимального развития за несколько дней до рождения. У такого эмбриона обнаруживаются уже вполне адекватные, и главное, видоизменяющиеся рефлекторные ответы на тактильные раздражения: прикосновение волоском к коже около уха вызывает специфическое подергивание последнего, непрерывное продолжение этого раздражения или его многократное повторение – приведение конечности данной стороны к раздражителю, чем нередко достигается его удаление; если же и после этого продолжать раздражение, то приходит в движение вся голова, а затем и туловище эмбриона, что может привести к его вращению, и наконец, по выражению Кармайкла, «каждая мышца приводится в действие». Кармайкл говорил по этому поводу, что эмбрион ведет себя как бы согласно пословице: «если не сразу достиг успеха – пробуй, пробуй еще», но подчеркивал, что нет оснований предполагать, чтобы что-либо в этом поведении являлось выученным.
Эмбриональное развитие поведения млекопитающих существенно отличается от такового у других животных. Это отличие выражается в том, что у млекопитающих движения конечностей формируются не из первоначальных общих движений всего зародыша, как мы это видели у вышеупомянутых других позвоночных, особенно низших, а появляются одновременно с этими движениями или даже раньше их. Вероятно, в эмбриогенезе млекопитающих большее значение приобрела ранняя афферентация, чем спонтанная эндогенная нейростимуляция.
Постоянная тесная связь развивающегося зародыша с материнским организмом, в частности посредством специального органа – плаценты, создает у млекопитающих совершенно особые условия для развития эмбрионального поведения. Новым и весьма важным фактором является в этом отношении возможность воздействия на этот процесс со стороны материнского организма, прежде всего гуморальным путем.
На такую возможность косвенно указывают результаты экспериментов, при которых на женских зародышей морской свинки еще во время их внутриутробного развития воздействовали мужским половым гормоном (тестостероном). В результате, став половозрелыми, они проявили признаки самцового поведения в ущерб сексуальному поведению, свойственному нормальным самкам. Аналогичное воздействие, произведенное после рождения, не давало такого эффекта. Подобным образом удавалось преобразовывать и половое поведение мужских особей. Очевидно, в ходе эмбриогенеза содержание тестостерона в организме зародыша влияет на формирование центральных нервных структур, регулирующих сексуальное поведение: его отсутствие – в сторону женских признаков, его наличие – в сторону мужских.
В экспериментах ряда исследований у беременных самок крыс периодически вызывали состояния беспокойства. В таких условиях родились более пугливые и возбудимые детеныши, чем в норме, несмотря на то, что их затем вскармливали другие самки, не подвергавшиеся экспериментальным воздействиям. Эти данные особенно отчетливо показывают роль влияния материнского организма на формирование признаков поведения детеныша в эмбриональном периоде его развития.
Пренатальное развитие сенсорных способностей и элементов общения
Влияние сенсорной стимуляции на двигательную активность эмбриона
Выше приводились примеры рефлекторных движений зародышей, производимых преимущественно в ответ на тактильные раздражения. Сенсомоторная активность составляет единый процесс на всех этапах жизни животного, хотя, как мы видели, двигательный компонент является в эмбриогенезе первичным и может возникнуть на эндогенной основе. Вместе с тем, по мере развития эмбриона и формирования его рецепторных систем, все большее значение приобретает сенсорная стимуляция, выступающая, очевидно, также в форме самостимуляции.
Куо видел такую самостимуляцию, в частности, в том, что куриный зародыш прикасается одной частью тела (например, ногой или крылом) к другой части (например, голове) и вызывает тем самым двигательную реакцию последней. Оппенгейм, правда ссылаясь на собственные исследования и работы других авторов, ставит под сомнение правомерность выводов Куо о таком механизме самостимуляции, но не отрицает существования эмбриональных сенсомоторных связей, как и значения сенсорной стимуляции в эмбриональном поведении.
Еще в начале 30-х годов Д.В.Орр и В.Ф.Уиндл сумели показать, что наряду со спонтанной двигательной активностью у куриного эмбриона развивается рефлекторная система движений. Изолированные движения крыла возникают в ответ на тактильное раздражение уже на ранних стадиях эмбриогенеза; это указывает на то, что потенциальные возможности рефлекторных реакций существуют уже тогда, когда еще отсутствуют реальные возможности внешней аффектации и двигательная активность зародыша проявляется лишь в общих спонтанных телодвижениях. Эти же ученые установили, что у куриного эмбриона моторные структуры нервной системы формируются раньше сенсорных, а первые реакции на внешние раздражения появляются лишь через четыре дня после первых спонтанных движений.
Однако наибольшее значение сенсорная стимуляция приобретает у куриного зародыша на последних стадиях эмбриогенеза, за 3–4 дня до вылупления (Гамбургер). Именно в этот период в развитие поведения включаются у птиц как мощные внешние факторы оптические и акустические стимулы, подготавливающие птенцов к биологически адекватному общению с родительскими особями.
Развитие зрения и слуха у эмбрионов птиц
Зрение и слух появляются лишь к концу эмбриогенеза и не влияют на развитие ранней двигательной активности зародыша. Правда, как было установлено рядом советских исследователей (Т. П. Блинкова, Г. Е. Свидерская и др.), сильные внешние раздражения способны вызвать реакции куриного зародыша уже на средних и даже ранних стадиях эмбриогенеза. Реакции на громкие звуки обнаруживаются не только после 14–19-го дня, когда уже начинает функционировать орган слуха, но и даже начиная с 5-го дня инкубации. В это же время можно вызвать реакции и на мощные световые воздействия. Все эти реакции выражаются в усилении или торможении эмбриональных движений. Однако, не говоря уже о том, что в данных экспериментах зародыши подвергались экстремальным, биологически неадекватным воздействиям, свет и звук могут на этом этапе выступать лишь как физические агенты, непосредственно влияющие на мышечную ткань или кожу, но не как носители оптической или акустической информации.
Если же, как явствует из новых данных американского ученого Г. Готтлиба, воздействовать на зародыш биологически адекватными, т.е. обычно встречающимися в природе, звуками на такой стадии, когда он еще не реагирует на подобные раздражения, то это может положительно сказаться на появляющихся позже слуховых реакциях эмбриона.
Что касается развития оптических реакций, то только начиная с 17–18-х суток инкубации в глазу и зрительных долях куриного эмбриона обнаруживаются электрофизиологические изменения в ответ на оптические раздражения. У зародыша пекинской утки, например, зрачковый рефлекс появляется на 16-й день инкубации, но это чисто фотохимическая реакция, которая не имеет функционального значения и сменяется на 18-й день (т.е. относительно раньше, чем у куриного эмбриона) подлинно нервной реакцией. Очевидно, к этому времени уже функционируют периферические и центрально-нервные элементы зрительного анализатора. Перед вылуплением птенцов зрачковый рефлекс практически уже так же развит, как у взрослой утки.
Развитие акустического контакта между эмбрионами и родительскими особями у птиц
У эмбрионов многих птиц в последние дни перед вылуплением не только начинают вполне функционировать дистантные рецепторы, т.е. органы зрения и слуха, но и появляются первые активные действия, направленные на внешнюю среду, а именно подача сигналов насиживающим родительским особям. Так, например, у представителя отряда чистиков кайры птенец научается еще за 3–4 дня до вылупления отличать голос родительской особи от голосов других кайр, гнездящихся в тесном соседстве на птичьих базарах. Если передискусственно инкубируемыми яйцами проигрывать магнитофонную запись криков какой-либо определенной взрослой кайры, а затем воспроизводить эту запись одновременно с записью криков другой кайры перед вылупившимися из этих яиц птенцами, то они направятся в сторону звуков, которые слышали еще до вылупления. Контрольные же птенцы из «неозвученных» яиц направятся в промежуток между источниками звуков, а затем начнут метаться между ними. Было установлено, что распознавание родительского голоса (в отличие от голосов соседних птиц) осуществляется на основе согласования ритмов подачи звуков родителя и невылупившегося птенца: в ответ на писк последнего насиживающая птица приподнимается, передвигает яйцо и сама подает голос. Таким образом, кинестетические ощущения эмбриона сочетаются с акустическими, а в целом его активность совпадает с таковой взрослой птицы, что и позволяет установить принадлежность услышанного звука родительской особи (исследования Б. Чанц).
В этом примере мы видим, как созревшее в эмбриогенезе врожденное, инстинктивное поведение (локомоторная реакция вылупившихся в изоляции птенцов на видоспецифический звук, т.е. на ключевой раздражитель) сочетается с истинным эмбриональным научением (условнорефлекторным путем), результатом которого является индивидуальное опознавание еще в пренатальном периоде развития, дифференцировка индивидуальных отличий видотипичных звуков. Именно в этом направлении совершается достройка соответствующего врожденного пускового механизма птенца, его обогащение необходимыми дополнительными признаками путем научения (в данном случае пренатального). Мы еще встретимся с этим вопросом при рассмотрении процессов запечатления. Сейчас же нам важно отметить, что между эмбрионом и родительской особью происходит обмен сигналами и возникает первичная коммуникация.
Сходные результаты были получены и у других видов птиц, в том числе у близкой родственницы кайры гагарки. Пренатальное распознавание голосов родителей было установлено также у гусиных (канадская казарка), куликов (кулик-сорока) и представителей других отрядов птиц. В частности, сравнение поведения птенцов, выведенных в инкубаторе в условиях звуковой изоляции, с таковым птенцов, слышавших крики взрослых птиц своего вида незадолго до вылупления, показало существенное значение эмбрионального научения и у чаек. Немецкая исследовательница М.Импековен установила, что изменения в поведении взрослых чаек (Larus atricilla) при переходе от насиживания к уходу за птенцами в большой степени обусловливаются акустическими сигналами, подаваемыми птенцами перед выклевом. При таких звуках насиживающая птица начинает посматривать вниз, на яйца, попеременно вставать и садиться, перекатывать яйца, отряхиваться и издавать ответные крики. Эти реакции удалось вызвать, воспроизводя издаваемые птенцом звуки в «чистом виде», т.е. в магнитофонной записи. При их замедленном проигрывании, т.е. изменении физических параметров звуков, реакции родительской особи ослабевают и появляются реже. С другой стороны, Импековен удалось экспериментально доказать, что родительские крики, услышанные птенцами еще до вылупления, стимулируют клевательные движения последних, в том числе и постнатальное клевание клюва родительской особи, т.е. «попрошайничество». Таким образом, стимулирующее влияние родительских криков проявляется здесь как результат пренатального накопления опыта.
Эмбриогенез и развитие психического отражения
Как видно из изложенного, в эмбриогенезе происходит интенсивная подготовка к последующим, постнатальным этапам формирования поведения, а отчасти и само формирование элементов поведения новорожденного путем, с одной стороны, развития генетически обусловленных компонентов активности и, с другой стороны, накопления эмбрионального опыта. Как и в постнатальной жизни животного, эти две стороны единого процесса развития поведения – врожденное и приобретаемое – невозможно отрывать друг от друга и изучать вне их взаимосвязи, поэтому неверно рассматривать эмбриогенез поведения с точки зрения альтернативы: созревание врожденных элементов поведения или эмбриональное упражнение. В каждом случае речь может идти лишь о том, какой из этих компонентов превалирует или даже доминирует. И здесь сложным образом переплетаются воздействия различных факторов в разных соотношениях и сочетаниях, как это происходит и на постнатальном этапе развития.
При этом необходимо, естественно, учитывать те специфические особенности, которыми отличается пренатальное развитие поведения животных. Это касается прежде всего роли внешней среды в формировании пренатальной двигательной активности и психической деятельности.
Приведенные выше данные показывают, что развитие поведения в пренатальном периоде онтогенеза совершается у низших и высших животных неодинаково, хотя и обнаруживает ряд общих черт. Эти филогенетические различия обусловлены закономерностями эволюции эмбриогенеза, установленными прежде всего Северцовым, о чем уже говорилось выше. Но в целом можно сказать, что у всех животных – во всяком случае на ранних стадиях эмбриогенеза – прямые влияния внешней среды играют незначительную роль (или не играют вообще никакой роли) в формировании отдельных форм двигательной активности.
Средой, в которой развивается эмбрион млекопитающего, является материнский организм, который не только хранит и защищает его от неблагоприятных воздействий, но и непосредственно обеспечивает всю его жизнедеятельность. Поэтому утроба является средой обитания зародыша, на которую направлена его активность. Однако связь эмбриона с подлинным внешним миром, в котором протекает вся постнатальная жизнь развивающегося организма, осуществляется лишь опосредованно через организм матери и не может иметь существенного значения для развития психики млекопитающего во внутриутробном периоде его развития.
В отличие от млекопитающих и, вероятно, других живородящих животных при внеутробном развитии зародыш часто подвергается воздействиям различных агентов внешней среды. Однако, как уже отмечалось, экспериментальные исследования показали, что эти агенты едва ли могут непосредственно направлять развитие первичных форм двигательной активности эмбриона. Но если даже допустить обратное, а именно что такие воздействия в состоянии оказать прямое влияние на формирование этой активности, то такая связь по необходимости была бы односторонней, ибо сфера двигательной активности зародыша не простирается дальше яйцевых оболочек и не может вызвать физические изменения в окружающей среде. Но это означает, что отсутствует важнейший источник психической деятельности.
Другое обстоятельство, крайне лимитирующее возможности развития психики в эмбриональном периоде развития, – это однородность, постоянство и бедность компонентами среды, которая окружает зародыша как в яйце (птичьем или икринке), так и в утробе млекопитающего. Там ему практически «нечего отражать». Поэтому будет, очевидно, правильно сказать, что психика эмбриона – это психика в процессе ее становления. Эмбрион – это еще не полноценное животное, а формирующийся организм животного на начальном этапе своего развития. Животная жизнь невозможна без активного взаимодействия с внешней (т.е. постнатальной) средой, а как раз это взаимодействие еще отсутствует на эмбриональном этапе развития, по меньшей мере на его ранних стадиях. В ходе эмбриогенеза осуществляется лишь подготовка к этому взаимодействию.
На ранних стадиях эмбриогенеза формируются предпосылки, потенциальные возможности психического отражения, т.е. существуют только зачаточные формы элементов психики. Лишь по мере того как формируются органы и системы органов развивающегося организма и появляется необходимость установления и расширения связей с внешним миром, зарождается и развивается психическое отражение, которое является функцией этих структур и служит установлению этих связей.
Как было показано, это происходит в конце эмбриогенеза, во всяком случае у птиц, у которых наряду с весьма дифференцированной двигательной активностью (и на ее основе) появляется коммуникативное поведение, обеспечивающее установление контактов с внешним миром (конкретно – с родительской особью) еще до вылупления.
Во взаимосвязях между невылупившимся птенцом и родительской особью, в согласовании их поведения на протяжении последних дней инкубации проявляется уже достаточно сложная психическая деятельность эмбриона. Правда, это относится лишь к заключительной стадии пренатального онтогенеза. При этом необходимо также учесть, что такая психическая активность зародыша едва ли свойственна млекопитающим, у которых эмбриональное развитие является по сравнению с птицами относительно укороченным: детеныши рождаются на более ранних стадиях эмбриогенеза, на которых можно предположить наличие лишь примитивных элементов будущей психической активности.
Итак, можно сказать, что значение эмбриогенеза для формирования психической деятельности состоит в том, чтобы подготовить морфофункциональную основу психического отражения. Это относится как к двигательным компонентам психической деятельности, так и к подготовке условий для функционирования сенсонейромоторных систем на постэмбриональном этапе развития.
Ясно, что будет поздно, если эти предпосылки начнут формироваться лишь после появления животного на свет, поэтому такая база должна уже существовать к началу постнатального развития, чтобы организм мог приступить к построению всесторонних отношений с компонентами среды его обитания. Для подготовки этой базы достаточна генетически фиксированная спонтанная двигательная активность эмбриона, дополняемая более или менее выраженным эмбриональным научением. Такая активность может формироваться и в той константной, однообразной, нерасчлененной, бедной предметными компонентами среде, в которой живет и развивается зародыш.
Глава 3 РАЗВИТИЕ ПСИХИЧЕСКОЙ ДЕЯТЕЛЬНОСТИ В РАННЕМ ПОСТНАТАЛЬНОМ ПЕРИОДЕ
Особенности постнатального развития поведения животных
Пре- и постнатальное развитие психической деятельности
Психическая деятельность эмбриона – это поведение и психика в процессе их становления на начальной стадии существования особи. Эмбрион является всего лишь формирующимся организмом, еще не способным к полноценному осуществлению функций, необходимых для установления жизненно важных взаимоотношений со средой обитания вида. В такой же степени и психика существует на пренатальной стадии развития лишь в зачаточной форме.
Рождение является поворотным пунктом, где весь процесс развития поведения получает новое направление. Естественно, что на уровне постнатального развития появляются совершенно новые факторы и закономерности, обусловленные взаимодействием организма с подлинной внешней средой. В этих новых условиях, однако, продолжается созревание врожденных элементов поведения и их слияние теперь уже с постнатальным, индивидуальным опытом. Поэтому, несмотря на коренные различия в условиях формирования пре- и постнатального поведения, между этими этапами онтогенеза поведения не только нет разрыва, но существует прямая преемственность. Именно в этом проявляется преадаптивное значение эмбрионального поведения, о чем также говорилось выше.
Если взять, к примеру, птиц, то, как было показано, к моменту вылупления уже существует достаточно развитая система общения между птенцом и высиживающей особью. Кроме того, существует акустическое общение и между невылупившимися птенцами внутри кладки, что влияет непосредственно на процесс вылупления. Так, у виргинской куропатки и японского перепела звуки, подаваемые одними птенцами, стимулируют активность других. В результате вылупление происходит дружно в течение приблизительно одного часа. Экспериментально этот эффект был доказан путем подкладывания яиц, находящихся на более ранней стадии инкубации, к яйцам, в которых находились птенцы перед самым вылуплением, что приводило к ускорению вылупления «отставших» птенцов на несколько часов. Акустическая коммуникация приводит, следовательно, у еще не вылупившихся птенцов к синхронизации вылупления. Преемственность же между эмбриональным и постэмбриональным поведением выражается в данном случае в том, что птенцы одного выводка, равно как птенцы и родители, отчасти уже «знают» друг друга как конкретных особей по индивидуальным признакам еще до появления первых на свет. Другие примеры, иллюстрирующие преадаптивное значение пренатального поведения, приводились в предыдущей главе.
Ранний постнатальный период имеет особенно большое значение для жизни особи, так как на этом этапе развития формируются важнейшие взаимоотношения организма с окружающей средой, устанавливаются связи с жизненно важными компонентами этой среды и закладываются основы поведения взрослого животного. Орбели говорил по этому поводу, что ранний постнатальный период является особо чувствительной фазой, наиболее жадно воспринимающей влияния, имеющие большое значение для дальнейшего формирования особи.
Разнообразие постэмбрионального развития поведения животных
Говоря о постнатальном развитии поведения животных, необходимо прежде всего иметь в виду, что оно протекает по-разному у разных животных и отличается специфическими закономерностями. Особенно это относится к тем животным, у которых новорожденные и ювенильные (т.е. молодые) формы резко отличаются строением и образом жизни от взрослых форм. У большинства беспозвоночных, а также ряда низших позвоночных личинки совершенно не похожи на взрослые особи, и зачастую им свойственны совершенно иные способы передвижения, питания и т.д.
Так, например, у морского моллюска из рода Strombus, как и у других морских брюхоногих, молодь ведет первоначально планктонный образ жизни, однако сразу же после опускания на дно и метаморфоза (т.е. превращения из одной стадии развития в другую) появляются свойственные взрослым животным формы придонной локомоции и питания. Вместе с тем защитная реакция – уход от хищника (хищных моллюсков рода Conus) – появляется только спустя три недели после метаморфоза, причем также сразу в «готовом виде». Предшествующий опыт (в эксперименте – контакт с водой, в которой содержался хищник) не ускоряет созревания этой реакции и не влияет на выраженность ее проявлений. В указанный же срок молодой Strombus тут же адекватно реагируют на приток воды, в которой находился его враг. Таким образом, мы имеем здесь дело со случаем весьма консервативного поведения, в той же мере генетически обусловленным, как и морфологические признаки. Постепенное усложнение поведения идет здесь строго «в ногу» с усложнением нервных структур и общего строения моллюска.
Аналогичные примеры известны также по членистоногим и другим беспозвоночным. Во всех этих случаях постэмбриональное развитие поведения совершается особенно сложным путем, ибо происходит резкая смена форм поведения и как бы в недрах личиночного поведения созревает качественно отличное взрослое поведение. Так, в приведенном примере личинка Strombus плавает на протяжении первой постэмбриональной стадии – в составе планктона – в поверхностных слоях моря, а затем переходит к придонному образу жизни и передвигается по субстрату характерными «прыжками».
Однако и в тех случаях, когда между ювенильными и взрослыми животными не обнаруживаются столь глубокие различия, филогенетические и экологические детерминанты вносят большое разнообразие в протекание постнатального онтогенеза поведения. Особенно большое значение имеет при этом степень зрелорождения потомства.
Постэмбриональное развитие поведения у зрело- и незрелорождающихся позвоночных
Детеныши позвоночных рождаются на разных стадиях зрелости. В предыдущей главе мы уже познакомились с примером крайнего незрелорождения – кенгуренком, появляющимся на свет еще в «полузародышевом» состоянии. Незрелорожденными, хотя и на более поздней стадии развития, являются птенцы многих птиц (воробьиных, хищных и др.), детеныши большинства млекопитающих (грызунов, хищных и др.). С другой стороны, птенцы кур, уток, гусей и ряда других птиц, как и детеныши копытных, представляют собой пример зрелорождения. В этом случае новорожденный почти сразу же, во всяком случае очень скоро, становится способным к самостоятельному выполнению основных жизненных функций (опорно-локомоторных, пищедобывательных и др.). Незрелорождающиеся же детеныши на протяжении определенного времени выполнять эти функции еще не могут и всецело зависят от родительского ухода. Промптов говорил в этом случае о «раннем биостарте». Под биостартом он понимал начало влияния биологических факторов на развитие птенца. Крайние проявления зрелорождения встречаются, правда, сравнительно редко. Чаще всего наблюдаются разные переходные степени зрелорождения. Например, у турухтана (отряд кулики) вылупившиеся птенцы проявляют много признаков зрелорождения как в рецепторной, так и в моторной сфере. В первые часы жизни они уже способны самостоятельно двигаться, затаиваться, вскоре они становятся способными и самостоятельно питаться. Вместе с тем они до 10-дневного возраста нуждаются в обогреве, как незрелорождающиеся птенцы, и координация движений вполне устанавливается лишь к 4-м суткам жизни. К этому сроку относится и полное формирование пищедобывательного поведения.
Орбели дал четкую биологическую характеристику развития поведения у зрело- и незрелорождающихся животных. Он указал на то, что, поскольку зрелорождающиеся детеныши начинают подвергаться воздействию среды в уже почти вполне сформированном состоянии, они сравнительно мало подвержены вредным влияниям среды. Но при этом чрезвычайно ограничены возможности дальнейшего прогрессивного развития поведения: имеют место лишь отдельные дополнительные надстройки условнорефлекторной деятельности.
Совершенно иное положение у животных, у которых развитие даже наследственно фиксированных, врожденных форм поведения выносится далеко за пределы внутрияйцевого или внутриматочного периода, т.е. у незрелорождающихся. По Орбели, эти детеныши, хотя им приходится труднее и они не могут жить без родительской помощи, находятся все же в более выгодном положении: развитие нервной системы у них еще не вполне закончено, и они могут подставить свои еще развивающиеся врожденные формы поведения под влияние агентов окружающей среды. В результате эти формы поведения в значительной степени модифицируются на основе переплетения врожденных и приобретаемых компонентов в соответствии с конкретными условиями среды. Как пишет Орбели, эти животные «родятся с настолько еще малосформированной нервной системой, что все дальнейшее постнатальное развитие представляет собою сплошную переработку наследственных форм и вновь возникающих условных форм поведения».*
* Орбели Л.А. Вопросы высшей нервной деятельности. М.;Л., 1949. С. 356.
С этой точки зрения более прогрессивными следует, безусловно, считать незрелорождающиеся виды животных, так как им начиная уже с «биостарта» обеспечено более пластичное, лабильное поведение, т.е. большая приспособляемость к постоянно меняющимся условиям среды. Правда, к кенгуру это не относится, так как здесь имеется, по существу, лишь кратковременный перерыв эмбриогенеза, как бы переход из одной утробы в другую, и ранняя смена способа питания – ведь в сумке матери крайне недоразвитый детеныш также весьма изолирован от внешнего мира и защищен от его вредных воздействий. (Важное отличие составляет, правда, возможное нахождение в той же сумке старшего детеныша.)
В этой связи интересно отметить, что психически наиболее развитые животные (а также человек, как подчеркивает и Орбели) в большинстве относятся к незрелорождающимся. У них новорожденный очень рано попадает в «настоящую» внешнюю среду, и генетически фиксированные формы поведения претерпевают здесь глубокую и длительную переработку. Правда, это не единственное обстоятельство, определяющее высокий уровень психического развития, ибо, например, гуси или лошади, т.е. типично зрелорождающиеся животные, также обнаруживают этот уровень.
По-видимому, решающее значение здесь имеют генетически обусловленные возможности накопления индивидуального опыта и его переноса в новые ситуации, активные воздействия на окружающую среду на основе развития эффекторно-рецепторных систем, двигательной активности, а в этом отношении различия между зрело- и незрелорождающимися животными сводятся к различиям в сроках формирования и развертывания этих систем и активности.
Что же касается этих сроков, т.е. общей скорости постнатального развития поведения, то они не отражаются на его результатах. Продолжительное постнатальное созревание, достройка и перестройка врожденного поведения, не влечет за собой замедления, задержки или тем более застоя общего развития поведения и наоборот. Японский орнитолог Н. Курода сопоставлял у разных видов воробьиных продолжительность гнездового периода и соответствующих форм гнездового поведения со сроками наступления способности к самостоятельному добыванию пищи у молодых птиц. Оказалось, что эта способность появляется приблизительно в одинаковые сроки независимо от длительности гнездового периода. Таким образом, при удлинении или укорочении гнездового периода птенец ничего не теряет, а в первом случае он быстро наверстывает «упущенное время» и оказывается не менее подготовленным к самостоятельной жизни, чем птицы, которые раньше покидают гнездо. Если учесть, что между зрело- и незрелорождением существуют переходные формы, то становится понятным, что и отмеченные различия имеют лишь относительное значение, тем более что темп онтогенеза на разных этапах меняется.
Значение заботы о потомстве
Большое значение, особенно у незрелорождающихся животных, приобретает родительская забота о потомстве, т.е. действия животных, обеспечивающие или улучшающие условия выживания и развития потомства. В процессе эволюции у многих групп животных возникли приспособления к защите и питанию развивающегося потомства со стороны родительской особи. Сюда относится и прохождение эмбриональных стадий развития в теле матери. Однако понятие «забота о потомстве» применяется лишь к постэмбриональному периоду. В ряде случаев забота о потомстве ограничивается созданием убежища и заготовкой пищи для будущего потомства, но материнская особь при этом не встречается с ним (превентивная забота о потомстве). Так, некоторые осы откладывают яйца на парализованных ими насекомых, которых прячут в специально вырытых норках, но затем уже не заботятся о вылупившихся личинках.
Более высокой формой заботы о потомстве является уход за потомством, проявляющийся в двух основных формах: пассивной и активной. В первом случае взрослые особи носят с собой яйца или молодых животных в специальных кожных углублениях, складках, сумках. Молодые животные при этом иногда питаются выделениями материнской особи. Эта форма ухода за потомством встречается у отдельных видов иглокожих, ракообразных, моллюсков, пауков, рыб (морской конек и игла, некоторые тропические окунеобразные – цихлиды), земноводных (жаба-повитуха, американская пипа, лягушка gastrotueca marsupiata), низших млекопитающих (ехидна, сумчатые). При активном же уходе за потомством взрослые особи выполняют специфические действия, направленные на обеспечение всех или многих сфер его жизнедеятельности – личинок насекомых, молоди рыб, птенцов, детенышей млекопитающих. Кроме устройства убежищ, кормления, обогрева, защиты, очищения поверхности тела и т.п. родители у многих высших животных (птиц и млекопитающих) также обучают свое потомство (например, находить пищу, распознавать врагов и т.д.).
Именно активный уход за потомством, высокоразвитая забота о нем делают возможным незрелорождение, а тем самым и все обусловленные им особенности психического развития. При этом эволюция заботы о потомстве знаменовалась, с одной стороны, интенсификацией и дифференциацией действий родителей по отношению к потомству, с другой стороны, усилением его зависимости от взрослых животных. Одновременно резко понизилась плодовитость. Однако возрастающая забота о потомстве влечет за собой и растущее противоречие между потребностями родительской особи и ее потомства. Это противоречие регулируется естественным отбором в сторону наибольшего прогресса вида. В.А.Вагнер охарактеризовал это формулой: минимум жертв матери – максимум требований потомства.
Таким образом, прогрессивные эволюционные приобретения, обеспечившие более гибкое приспособление растущего организма к условиям его жизни в постнатальном онтогенезе, имеют весьма сложную природу и включают в себя разные формы заботы о потомстве в зависимости от степени зрелорождаемости. Всем комплексом этих факторов определяется в каждом случае конкретный ход постнатального развития поведения.
Инстинктивное поведение в раннем постнатальном периоде
Инстинктивные движения
Стабильные, жесткие инстинктивные движения появляются в раннем постнатальном онтогенезе в настолько «готовом» виде, что долгое время считалось, что они совершенно не развиваются и не нуждаются в индивидуальном упражнении. На самом деле, как мы уже знаем, вполне врожденной является лишь «программа» их формирования в ходе онтогенеза. Сам же этот процесс обусловлен развитием экзосоматических эффекторных органов (и соответственных центральнонервных структур), функцией которых инстинктивные движения и являются. Только в таком смысле можно говорить о не зависящем от влияний среды появлении или созревании инстинктивных движений в онтогенезе.
Поскольку эти вопросы уже обсуждались раньше, мы ограничимся здесь лишь несколькими дополнительными замечаниями. Формирование инстинктивных движений в онтогенезе выявляется экспериментально путем изолированного выращивания детеныша с момента рождения (по методу «Каспар Гаузер»). Опыты, проведенные на птицах и млекопитающих (преимущественно грызунах), показали, что хотя поведенческие акты таких животных обнаруживают существенные отклонения от нормы, моторные элементы выполняемых при этом движений остаются без изменений, несмотря на то что детеныши были лишены возможности упражнения. (Курода, например, определил у птенцов серого скворца около 50 подобных элементов поведения, появляющихся спонтанно в разные сроки раннего постэмбрионального развития.) Это относится к особенно важным для жизни движениям, связанным с локомоцией, едой, спариванием, комфортными действиями и т.д. Но речь идет только о двигательных элементах, из которых слагаются эти движения. Длительность, частота, направленность, координированность, «отработанность» и совершенство этих движений, не говоря об их согласованности с другими поведенческими актами, а тем более с действиями других животных, оказывались в большей или меньшей степени нарушенными.
Большую известность приобрели проведенные американским ученым Э. Гессом опыты с цыплятами, которые вылуплялись в темноте, а затем были снабжены призматическими очками, смещавшими поле зрения на 7 градусов вправо. На 4-й день сопоставление с контрольными птенцами (снабженными очками, не искажавшими зрение) показало, что цыплята обеих групп клевали по целевому объекту (шляпке гвоздя) более точно, чем в первый день, но при этом не изменилось общее направление ударов: контрольные животные клевали по-прежнему на 7 градусов вправо от цели. Это доказывает, что данная двигательная координация, клевание и общее направление этих движений в поле зрения являются врожденными и не зависят от индивидуального опыта. Последний, однако, необходим для «отработки» точности ударов по цели, для обеспечения эффективности этой реакции.
Аналогично обстоит дело и у млекопитающих. Даже у обезьян было установлено, что моторные элементы двигательных реакций развиваются у новорожденных и без афферентной обратной связи (в эксперименте над макаками и павианами – после деафферентации передних конечностей у ослепленных детенышей). Вместе с тем, как показали, например, американский исследователь Г. Харлоу и его сотрудники, у макак-резусов и шимпанзе, выращенных в изоляции, появляются некоторые двигательные элементы нормального полового поведения, однако эти элементы не интегрируются в единый поведенческий акт, а в результате сексуальное поведение этих животных становится ненормальным. Аналогичные данные были получены и у морских свинок.
Таким образом, постнатальное развитие инстинктивных движений обнаруживает более или менее четкую генетическую обусловленность, не зависящую от раннего опыта. Но это относится только к элементарным врожденным двигательным координациям, но не к целым инстинктивным двигательным актам.
Врожденное узнавание
Появившись на свет, животное должно быстро и верно ориентировать свои движения по отношению к разным агентам среды. Такая ориентация по отношению к жизненно важным компонентам среды с минимальной тратой энергии и времени – поистине вопрос жизни и смерти, особенно для зрелорождающихся животных. Осуществляется она в первую очередь путем так называемого врожденного узнавания.
Врожденное узнавание (по Фабри) проявляется во врожденном, не зависящем от индивидуального опыта видоспецифическом избирательном отношении животных к определенным компонентам окружающей среды, признакам объектов, ситуациям, в способности животных биологически адекватно реагировать на некоторые признаки еще незнакомых им объектов или ситуаций. Таким образом, мы имеем здесь дело с врожденной формой ориентации, проявляющейся в полезных для особи (и вида) реакциях на признаки существенных компонентов среды без предварительного научения, с проявлениями «видовой памяти».
Подобные видотипичные положительные или отрицательные реакции животных, еще не успевших накопить индивидуальный опыт, направлены на такие компоненты среды, как, например, пища, материнская особь, особи своего вида; враги (или ситуации тревоги, даже если враг непосредственно не воспринимается) и т.п. Словом, речь идет о некоем преадаптивном узнавании объектов видотипичного инстинктивного поведения, позволяющего животному опознать биологически значимые объекты по некоторым их элементарным специфическим признакам без предварительного научения.
Основу такого опознавания составляют таксисы, но далеко не всегда здесь действуют простые безусловные рефлексы. Как показал Лоренц, во многих случаях подобная специфическая направленность поведения животного основана на соответствующих врожденных пусковых механизмах; сами же признаки, ориентирующие поведение животного, представляют собой ключевые раздражители.
Таким образом, оказавшись в биологически значимой ситуации, животное как бы узнает то, что никогда еще не воспринималось, с чем еще не приходилось познакомиться. Имеет место предварительная готовность к встрече с самыми важными для выживания элементами среды, т.е. как бы предваряется возможный будущий опыт особи.
Примеров врожденного узнавания можно привести множество. Напомним вновь о поведении новорожденного кенгуренка – его безошибочном нахождении сумки по ряду признаков, с которыми ему до этого не приходилось сталкиваться и в распознавании которых он не мог упражняться. Птенцы дроздов уже в первый день после вылупления, еще будучи слепыми, узнают о прилете родителей (с кормом) по легкому сотрясению гнезда и отвечают на это вытягиванием шеи и раскрытием клюва. Птенцы зреловылупляющихся птиц реагируют на вид подвижного объекта над головой припаданием к земле, что является эффективной защитой от воздушных врагов – хищных птиц. Крольчата в первый же день после рождения ориентируются в пространстве по тактильным раздражениям и таким образом находят источник пищи – соски матери. На каждое легкое прикосновение они реагируют движением в сторону раздражителя: при раздражении переднего конца головы перемещаются вперед, при прикосновении к спине или хвосту поворачиваются назад. Поворотами в сторону прикосновения реагируют также щенки и другие новорожденные млекопитающие.
Во всех случаях врожденного узнавания имеет место реакция на отдельные простые признаки биологически значимых объектов или же на сочетания или соотношения таких признаков, что закономерно и для реакции на ключевые стимулы. Имеет место и специфическая врожденная готовность к ощущению этих стимулов, характерная для врожденных пусковых механизмов. Всем этим и определяется избирательность врожденного узнавания, ее направленность лишь на определенные компоненты среды.
Эта отличительная особенность врожденного узнавания, как и вообще все связанные с ним закономерности, по Фабри, непосредственно обусловлена развитием врожденной двигательной активности. Как всегда, психическое развитие совершается и здесь в ходе и в результате развития моторики: врожденное узнавание возможно лишь как результат определенного уровня и специфической направленности развития врожденных компонентов моторики, о которых только что говорилось, т.е. тех специфических элементов движений, из которых слагается целостная реакция животного. При этом важно подчеркнуть, что речь идет именно о врожденных компонентах двигательной активности, появляющихся (или созревающих) в ходе раннего постнатального онтогенеза без научения.
Поясним сказанное примером. В лаборатории А. Д. Слонима осуществлялось с первого дня жизни перекрестное воспитание детенышей двух видов грызунов: большой песчанки (роющая форма) и белой крысы (нероющая форма). Независимо от этих условий молодые песчанки с 17-го дня жизни начинали проявлять типичную для их вида роющую деятельность, в то время как у крысят, которые воспитывались песчанками, роющие движения намечались в весьма слабом виде и то лишь начиная с 52-го дня жизни.
О чем говорят результаты этих опытов? Совершенно очевидно, что лишь после того как сформировались соответствующие двигательные способности конечностей, т.е. после того, как независимо от внешних условий воспитания появились врожденные, инстинктивные роющие движения, наступило врожденное инстинктивное узнавание рыхлого материала (песка, который ежедневно предъявлялся наряду с деревянными палочками в течение 10–15 минут) как возможного объекта рытья. Следовательно, избирательное узнавание и выбор этого материала по признаку рыхлости (ключевой раздражитель!) обусловлены необходимостью выполнения определенных движений, осуществления одной из жизненно важных для данного вида функций конечностей.
Врожденное узнавание и ранний опыт
В начале постнатального развития животного недопустима «роскошь» продолжительного научения. Тем не менее уже здесь проявляется общее правило, что нет чисто врожденных форм поведения, лишенных каких бы то ни было элементов научения. Это всецело относится и к врожденному узнаванию, которое всегда обогащается, корректируется или перестраивается в результате приобретения раннего опыта, проявляющегося в разных формах постнатального научения.
Так, нередко встречаются явления привыкания к часто повторяющимся биологически малозначащим стимулам. Птенцы куриных птиц, первоначально одинаково реагирующие на каждый подвижный объект, появляющийся над ними, вскоре научаются распознавать безопасные объекты (рис. 20). Это происходит путем привыкания: после неоднократного притаивания в ответ на появление в поле зрения безобидного объекта (листа, безопасной птицы и т.п.) эта реакция ослабевает и наконец совсем исчезает. Однако привыкает птенец лишь к объектам, которые он видит часто. В отношении же редких объектов необычного вида (а к таким и относятся хищные птицы с их характерным силуэтом) реакция, основанная на врожденном узнавании подвижного объекта в воздухе над головой, а именно затаивание, сохраняется в прежнем виде. Таким образом, мы имеем здесь дело с уточнением, дифференцировкой врожденного узнавания путем приобретения раннего опыта. Результатом этого процесса является в данном случае дифференцированное распознавание безопасных объектов.
Сходным образом, как уже указывалось, обстоит дело и у рыб, мальки которых на основе врожденного узнавания первоначально уплывают от любого крупного объекта или тени, надвигающихся на них. Впоследствии же они перестают реагировать на безопасные объекты, но научаются распознавать отличительные признаки хищника путем наблюдения за поимкой и поеданием членов стаи.

Рис. 20. Привыкание у птенца (по Тинбергену). Пояснения см. в тексте
В результате раннего научения может меняться и сигнальное значение раздражителей. Молодь осетровых рыб реагирует первоначально на свет отрицательно, но начиная с 5-го дня (у шипа) или 9-го дня (у осетра) эта реакция превращается в положительную под влиянием перехода к активному питанию и образованию соответствующих пищевых условных рефлексов. В последующие дни реакция мальков на свет может индивидуально меняться в зависимости от конкретных условий кормления (В.Ю.Касимов).
В других случаях происходит достройка или изменение врожденного узнавания путем включения новых сенсорных систем. Так, дроздята, как уже упоминалось, в первые дни после вылупления реагируют на сотрясение гнезда выгягиванием шеи и раскрытием клюва. Пространственная ориентация этой реакции осуществляется на основе гравитационной чувствительности, локализованной в органе равновесия во внутреннем ухе, а шея вытягивается вертикально вверх независимо от места расположения источника раздражения. Эта врожденная реакция сохраняется и после прозрения птенцов, в недельном возрасте, хотя теперь уже она осуществляется и на зрительный стимул – появление какого-либо объекта в поле зрения птенца. Только постепенно, по прошествии еще нескольких дней, зрение становится ведущей рецепцией, и в конце концов дроздята научаются четко локализовывать оптический стимул, каковым в норме является родительская особь с кормом, и поворачивать голову в сторону его появления.
У молоди осетровых рыб в течение 1–3 дней после выклева, т.е. до перехода к активному питанию, появляется реотаксисная реакция (ориентация тела по потоку воды), которая в отличие от других рыб сопряжена с тигмотаксисом (ориентация по тактильным раздражениям, обусловленным прикосновениями к посторонним предметам): реотаксис проявляется в основном лишь при контакте малька с субстратом, что препятствует его смыванию течением. Только после появления положительно-фототаксисной реакции тактильная ориентация сменяется оптической (оптомоторной реакцией) и малек становится способным ориентировать свое тело и в толще воды (Д.С.Павлов). Это включение новых сенсорных систем, модификация врожденного узнавания, также происходит в условиях накопления первичного сенсомоторного опыта.
Рассматривая взаимоотношения между врожденным узнаванием (и врожденным поведением вообще) и ранним научением, необходимо постоянно помнить о том, что характер этого взаимоотношения в каждом конкретном случае определяется экологическими особенностями данного вида. Как в количественном, так и в качественном отношении эти взаимоотношения будут существенно различаться, например, у рыб, обитающих в различных условиях освещения, температуры, движения воды и т.д.
Так, у пресноводных рыб, нерестящихся в проточной холодной воде на значительной глубине (сиг), личиночный период короткий, личинки сразу начинают активно плавать, и отрицательный геотаксис, а затем и положительный фототаксис приводят их в поверхностные слои воды. Личинки же рыб, нерестящихся в мелководных, слабопроточных, поросших растительностью водоемах (сазан, щука), долго пребывают на месте, приклеиваясь (выделениями специальных желез) к растениям. Решающее значение в этом смысле имеет врожденное узнавание на основе тигмотаксиса, обеспечивающего тесный контакт животного с субстратом. К поверхности воды эти мальки поднимаются по мере своего развития лишь постепенно, как бы по ступенькам, причем тигмотаксис все больше слабеет, положительный же фототаксис все усиливается. Понятно, что и условия накопления первичного опыта, а тем самым и взаимоотношения между врожденным узнаванием и ранним научением, будут у этих двух групп рыб в корне различными.
Ранний опыт
Облигатное научение
Приведенные выше примеры постнатального научения относятся к упомянутому ранее облигатному научению. Сюда относятся все формы научения, которые в естественных условиях совершенно необходимы для выполнения важнейших жизненных функций, т.е. относящихся к видоспецифическому, инстинктивному поведению. Результаты облигатного научения в одинаковой мере необходимы для выживания всем представителям данного вида, и эта видоспецифичность сближает облигатное научение с врожденными формами поведения. Облигатное научение и врожденное поведение, в частности врожденное узнавание, теснейшим образом связаны друг с другом в единый комплекс. В этом единстве и находит свое воплощение модификация врожденного поведения ранним опытом. Не только сама способность к облигатному научению, но и конкретные ее проявления генетически фиксированы в такой же степени, как и форма инстинктивного поведения.
Характерным признаком облигатного научения является также то, что оно может осуществляться только на протяжении определенных, так называемых сенсибильных (или чувствительных), периодов онтогенеза. Ни до, ни после этого облигатное научение невозможно. Как еще будет показано, эти периоды начинаются чаще всего очень скоро после появления животного на свет и являются непродолжительными. В отдельных случаях, правда, они приходятся на более поздние сроки и могут обнаруживаться даже у взрослых животных, равно как могут быть и относительно длительными.
В отличие от облигатного научения факультативное научение представляет собой, как нам также уже известно, приобретение индивидуального опыта, который зависит от частных условий жизни особи и не является необходимым для всех представителей данного вида в качестве компонента их инстинктивного поведения. Факультативное научение модифицирует, совершенствует и приспособляет видотипичное, врожденное поведение в соответствии с особыми, частными, преходящими, а зачастую и случайными элементами среды обитания особи, поэтому факультативное научение носит сугубо индивидуальный характер, оно не приурочено к определенным сенсибильным периодам и отличается большой лабильностью и обратимостью. Видоспецифичными являются здесь лишь сама способность к научению и пределы этой способности.
Между облигатным и факультативным научением не всегда возможно провести резкую грань, и в реальном поведении животных, особенно молодых, зачастую обнаруживаются сочетания этих двух категорий приобретения индивидуального опыта (см. ниже). Наиболее четко и часто факультативное научение проявляется в виде разного рода навыков, которые уже рассматривались раньше. Что же касается привыкания, то оно, как было показано, может встречаться и в сфере облигатного научения.
Помимо уже отмеченных сфер поведения облигатное научение важно и для формирования пищевого поведения.
Это проявляется прежде всего в том, что путем облигатного научения детеныши во многих случаях узнают отличительные признаки пищевых объектов. Например, экспериментально доказано, что первоначальный прием пищи служит у змеенышей источником накопления опыта, определяющим последующее распознавание пищевых объектов по их химическим признакам. Детенышам морских свинок предъявляли в течение первых девяти дней после рождения различные съедобные и несъедобные объекты, в результате чего и формировалось предпочтение съедобных объектов. Распознавание несъедобных объектов без такого предварительного контакта оказалось невозможным. Это соответствует тому, что в школе Павлова изучалось на щенках (распознавание вкуса и запаха мяса) и получило название «натуральный условный рефлекс».
Отметим в этой связи, что Г. И. Поляков считает такие рефлексы низшей и наиболее древней категорией условнорефлекторной деятельности, функцией старой коры головного мозга и поэтому особенно характерными для пресмыкающихся, у которых условные рефлексы на неестественные, искусственные стимулы вырабатываются лишь с трудом. У млекопитающих же, по Полякову, натуральные условные рефлексы встречаются, как правило, только в раннем постнатальном онтогенезе, когда новая кора еще не созрела. Именно таким путем, например, котенок научается распознавать писк мыши.
Не только типичные признаки пищевого объекта усваиваются детенышами путем облигатного научения. Важное значение это научение имеет и для формирования пищедобывательных приемов, т.е. двигательных актов, обеспечивающих захват и само потребление пищевых объектов, у хищников – овладевание и поедание добычи. Правда, в своей основе эти движения являются врожденными двигательными координациями и созревают независимо от индивидуального опыта, как и другие инстинктивные движения. Однако их достройка, совершенствование и модификация происходят в конкретных условиях постнатального онтогенеза в результате облигатного научения. Так, белка должна научиться наиболее эффективным образом вскрывать орехи, хотя необходимые для этого движения являются врожденными и уже присущи бельчонку в «готовом виде». Детеныши карликовой мангусты (Helogale undulata) оказываются к моменту выхода из гнезда (в 24-дневном возрасте) уже вполне способными к выполнению движений, необходимых для ловли добычи, хотя заниматься этим им еще не приходилось и они только облизывали и обсасывали убитых животных, которых приносили в гнездо родители. Но каждому детенышу приходится учиться правильно производить укус, мгновенно умерщвляющий добычу, а также учиться обращению с реже встречающимися видами жертв. Характерное для этих животных бросание пищевых объектов с твердой поверхностью (особенно яиц) под туловище, назад, осуществляется первоначально в несовершенном виде и несинхронно. И это действие совершенствуется путем научения.
Необходимо, однако, отметить, что подобная «отработка» инстинктивных компонентов поведения происходит у высших животных главным образом на следующем, игровом этапе постнатального развития поведения. Как будет еще показано, игра имеет в этом отношении исключительно большое значение для окончательного формирования поведенческих актов. У низших животных, у которых нет игровой активности и соответственно отсутствует второй этап постнатального развития поведения, облигатное научение выступает как единственный фактор постнатальной достройки инстинктивного поведения. Особенно это относится, очевидно, к беспозвоночным, хотя у этих животных облигатное научение, как и весь онтогенез поведения, еще совершенно недостаточно изучено. По имеющимся отрывочным данным можно, однако, заключить, что во всяком случае у насекомых облигатное научение может играть существенную роль в формировании поведения. Так, английский этолог В.Г.Торп и его сотрудники установили, что имагинальные, половозрелые формы насекомых ориентируются по запахам, влиянию которых они подвергались на определенных стадиях (сенсибильных периодах) их личиночного развития. Если, например, воздействовать на личинок плодовых мушек (дрозофил) запахом мяты, то они будут впоследствии, превратившись во взрослых насекомых, отыскивать для откладывания яиц субстрат с этим запахом. Наездник Nemeritis canescens паразитирует на личинках моли рода Ephestia, но если личинок наездника вырастить на личинках другого вида (Meliphora), который в естественных условиях не поражается этим наездником, то запах этого вида будет затем предпочитаться взрослыми насекомыми при поиске жертвы. Но этот эффект может быть достигнут лишь в том случае, если контакт с необычным хозяином (личинкой Meliphora) устанавливается в момент выхода личинок наездника из яйца или вскоре после этого. Только в этот краткий период запах нового хозяина оказывается действенным. Следовательно, опять же процесс научения приурочен к четко определенному сенсибильному периоду.
Однако запаховая ориентация насекомых определяется не единственно облигатным научением. Так, в указанном эксперименте у наездников сохранилось избирательное отношение к запаху естественного хозяина (Ephestia) наряду с благоприобретенной повышенной чувствительностью по отношению к запаху нового хозяина. Напрашивается вывод, что положительная хемотаксисная реакция в первом случае определяется врожденным узнаванием, во втором – облигатным научением, которое, как всегда, сочетается с этим инстинктивным компонентом поведения и в нормальных условиях подкрепляет и модифицирует его в соответствии с конкретными условиями развития особи.
Запечатление
Как уже отмечалось, ранний опыт слагается прежде всего из облигатного научения. Факультативное научение, если оно вообще происходит, здесь только дополняет, уточняет и конкретизирует процесс облигатного научения. Преобладание последнего в раннем постнатальном онтогенезе объясняется тем, что в этот период происходит достройка врожденных пусковых механизмов ряда важнейших инстинктивных действии путем включения в них индивидуально приобретаемых компонентов. Именно в этом состоит сущность процесса, получившего название запечатления (Pragung, Imprinting).
Запечатление является важным и характерным компонентом раннего постнатального онтогенеза. Это форма облигатного научения (с обязательным присутствием в качестве важных составных частей элементов факультативного научения), при котором очень быстро фиксируются в памяти отличительные признаки объектов инстинктивных поведенческих актов, поэтому запечатление квалифицируют и как «перцептивное научение», направленное на распознавание «незнакомого» в дополнение к «знакомому» (т.е. к врожденному узнаванию). Как и в других случаях облигатного научения, запечатление совершается лишь в течение определенных сенсибильных периодов, причем без пищевого или иного внешнего подкрепления. Результаты запечатления отличаются исключительной прочностью («необратимостью»).
Объектами запечатления являются родительские особи (выступающие одновременно как носители типичных признаков вида), братья и сестры (детеныши одного помета), будущие половые партнеры. Возможно также запечатление внешних признаков постоянных врагов, так как защитная реакция на эти признаки формируется при сочетании вида врага с предостерегающими криками родителей или, как выше отмечалось в отношении мальков рыб, при виде гибели сородича. При более широком понимании запечатления некоторые авторы относят к нему также формирование реакции на пищевые объекты, а также на характерные признаки места обитания (рождения), что, вероятно, не вполне правомерно, хотя процессы запечатления могут и сопутствовать формированию этих реакций. Здесь мы рассмотрим Запечатление лишь в его «классическом» понимании, а именно как связанное с явлениями общения.
Запечатление изучено преимущественно на зрелорождающихся животных, хотя оно свойственно и незрепорождающимся. Чаще всего запечатление происходит вскоре после рождения, в течение весьма ограниченного сенсибильного периода, нередко при первой же встрече с объектом запечатления. Не все ученые придерживаются одного мнения о сущности этого процесса, и некоторые из них (например, Гесс) склонны даже выделить запечатление в особую, отличную от научения категорию поведения. Однако большинство исследователей все же относят запечатление к научению. Особенно четко запечатление проявляется в реакции следования, изучению которой посвятили свои усилия многие ученые.
Реакция следования
Феномен этой реакции состоит в том, что зрелорождающиеся детеныши уже вскоре после появления на свет неотступно двигаются вслед за родителями (и одновременно друг за другом). Этот феномен общеизвестен по поведению детенышей домашних животных. Приведем несколько примеров реакции следования у диких животных.
Самка гоголя, небольшой утки, устраивающей свое гнездо в дуплах на высоте до 15 метров над землей, непосредственно перед вылуплением птенцов покидает гнездо. Она летит к водоему, откуда возвращается лишь спустя несколько часов, но обратно в дупло она уже не влетает, а начинает летать перед входом в дупло из стороны в сторону, издавая призывные крики. В ответ на это птенцы один за другим подскакивают к входному отверстию, откуда они, посмотрев на летящую перед ними птицу, бросаются вниз на землю. Приземлившись, они тут же поднимаются на ноги и начинают ходить.
Интересно отметить, что к подобным поразительным прыжкам способны и птенцы другого вида уток, которые, как правило, устраивают гнезда в норах, – пеганки. Иногда же эти утки гнездятся в местах, расположенных на высоте до 3–5 метров над землей. В этих случаях вылупившиеся утята прыгают, точнее, сваливаются вниз так же, как гоголята. Еще более «драматичные» прыжки совершают птенцы чистиковых, например гагарки, спрыгивающие с высоких крутых утесов, правда, уже в возрасте 19–20 дней.
Даже стукнувшись о прибрежную полосу суши, они тут же поднимаются на ноги и направляются к воде.
Вернемся, однако, к гоголятам. После того как весь выводок покинул описанным способом гнездо, мать, постепенно снижаясь, садится возле него, а затем, немного походив вокруг утят, направляется к водоему, до которого подчас до двух километров ходьбы. Утята сейчас же движутся вслед за матерью и друг за другом, развивая при этом поразительную скорость, а достигнув воды, они незамедлительно пускаются за матерью вплавь. (Сходным образом ведут себя и птенцы гагарок: мать, пролетев над птенцом, садится неподалеку на воду, и он направляется к ней.)
У млекопитающих реакция следования особенно хорошо выражена у копытных, у которых детеныши рождаются в весьма зрелом состоянии и очень быстро приобретают способность стоять и ходить. В связи с большой ролью, которую играет у этих животных обоняние, запечатление у них происходит не только на оптические и акустические, но и на ольфакторные признаки, т.е. на запахи родителей. Например, верблюжонок уже через 10 минут после рождения делает первые попытки подняться на ноги, а еще через 90 минут он уже может свободно стоять. В течение первых суток формируется и реакция следования за матерью. Вальтер описал формирование реакции следования у детенышей антилоп. У изолированных после рождения от матерей детенышей (в неволе) запечатление может произойти по отношению к проходящему мимо человеку, если детеныш увидит его во время сенситивного периода. Правда, советская исследовательница И. З. Плюснина считает, что привязанность к матери формируется у детенышей копытных (имеются в виду ягнята) не только на основе запечатления. Как мы еще увидим, это справедливо и для других животных, в частности для цыплят.
Среди грызунов реакция следования хорошо выражена, например, у детенышей морских свинок. В отличие от подавляющего большинства других грызунов морские свинки не устраивают гнезда, и детеныши рождаются у них вполне развитыми, зрячими. Вес новорожденного составляет 60–120 граммов. Он обладает уже постоянными зубами, поскольку смена молочных зубов происходит еще в утробе матери.
Описана реакция следования и у детенышей тюленей. Тюленята рождаются очень крупными – их величина достигает почти половины длины тела матери. Еще в ее утробе происходит линька и смена молочных зубов. Уже спустя 30 минут после появления на свет новорожденный (и даже извлеченный оперативным путем из тела матери) тюлененок в состоянии ползать подобно взрослым тюленям, плавать, нырять и вылезать на лед. При удалении матери ее детеныши немедленно двигаются вслед за ней. Время от времени мать стимулирует тюлененка, прикасаясь к нему мордой или с силой шлепая передними ластами по воде, обдавая его брызгами. Именно последнее служит «приглашением» к следованию.
Встречается реакция следования также у некоторых рыб, например у цихлид, о чем еще пойдет речь ниже.
Как видно из приведенных примеров, реакция следования ориентирована на родительскую особь (а также на других детенышей того же выводка). Она обеспечивает выживание детенышей, сплоченность животной семьи, возможность ее охраны со стороны взрослых особей и осуществления ухода за потомством. Без реакции следования родительская особь не могла бы осуществить заботу о потомстве, ибо для ее осуществления необходимо, чтобы детеныши в самые сжатые сроки научились отличать индивидуальные признаки своей матери, от которой нельзя отстать, что и достигается путем запечатления. Таким образом, быстрая конкретизация инстинктивного поведения детенышей на индивидуально опознаваемых объектах (родителях, собратьях) обеспечивает здесь формирование жизненно важных приспособительных реакций в максимально сжатые сроки.
Первые опыты по изучению реакции следования проводились еще в 70-х годах прошлого века Д.Сполдингом, но всестороннее изучение и теоретический анализ процессов запечатления, в том числе и реакции следования, были осуществлены Лоренцем в 30-х годах нашего века. С тех пор эти вопросы изучались многими исследователями, но незаслуженно забыто, что уже в то время опыты по изучению реакции следования ставились в лаборатории Боровского в Москве. В этих опытах перед цыплятами передвигалась черная коробка, и если ее «медленно двигали вдоль стола за привязанную к ней веревку, то большинство цыплят шли за ней... шли даже тогда, когда тем самым удалялись от той части стола, где был рассыпан корм».* Вместе с тем цыплята не следовали за белой коробкой и черным платком. Изолированные в момент вылупления, цыплята уже спустя сутки не реагировали на наседку, хотя та относилась к ним как обычно, но вместо этого следовали за рукой экспериментатора. В лаборатории Боровского ставились и другие опыты по этой теме, но затем лабораторное изучение реакции следования было в Советском Союзе возобновлено лишь в 60-х годах (исследования К.Э.Фабри и сотрудников). В ином плане запечатление (на неподвижный объект) изучалось впоследствии в лаборатории А. Д. Слонима.
* Боровский В.М. Поведение цыплят, выведенных в инкубаторе// Рефлексы, инстинкты, навыки. Т. 2. М., 1936. С.183.
Еще Сполдинг установил, что при реакции следования объектом запечатления, т.е. объектом видотипичных, инстинктивных действий, может быть не только любой представитель данного вида, но и другое животное или человек и даже неживой, но подвижный объект. Важно лишь, чтобы это был первый передвигающийся объект, увиденный в жизни. Цыплята, у которых была сформирована реакция следования за двигающимся шаром, не станут затем следовать за другим объектом или даже шаром, но иного цвета, а будут избегать его и неотступно двигаться вслед за «своим» шаром.
Как и в других случаях запечатления, реакция следования формируется в сжатые сроки: у кур и уток сенсибильный период начинается в первые же часы после вылупления, достигает оптимума через 10–15 часов, а затем постепенно сходит на нет. В приведенном выше примере выпрыгивание гоголят из дупла происходило именно в оптимальные часы сенсибильного периода, когда со времени вылупления еще не прошло 12 часов. У морских свинок сенсибильный период весьма растянут (с 6-го до 30–40-го дня жизни), но оптимальный срок – 6–7-й день. До 14-го дня «запечатляемость» еще сохраняется на довольно высоком уровне, но затем постепенно сходит на нет (И. З. Плюснина).
Уже упоминалось о том, что запечатление совершается с исключительной быстротой. Следовать за объектом запечатления птенцы начинают зачастую уже после первой встречи с ним, т.е. в эксперименте – после первого же показа. Но если даже и потребуется несколько предъявлений, реакция быстро достигает максимальной интенсивности и в дальнейшем уже остается на этом уровне до тех пор, пока птенцы не подрастут.
Большой интерес представляет вопрос о подкреплении реакции следования. Ведь детеныши следуют за объектом запечатления без всякого вознаграждения, а если искусственно затруднить следование или даже нанести птенцам болевые раздражения, то реакция следования становится только еще более интенсивной (Гесс). Объясняется это тем, что в отличие от факультативного научения облигатное научение не зависит от наличия или отсутствия не только случайных, маловажных компонентов среды, но и таких, которые могли бы служить для подкрепления результатов научения. Результаты облигатного научения настолько важны для жизни особи, что соответствующие реакции должны срочно формироваться при любых обстоятельствах, даже если в данный момент отсутствует возможность их внешнего подкрепления. Правда, при запечатлении имеет, очевидно, место «внутреннее» подкрепление по принципу обратной связи, источником которого являются ощущения от самих производимых движений, т.е. происходит лишь проприоцептивное подкрепление.
Отмеченная выше «необратимость» запечатления проявляется в чрезвычайной прочности реакции следования: практически объект запечатления не может в этом качестве заменяться другим объектом, и «реимпринтинг», т.е. переучивание на другой объект, как правило, возможен лишь в специальных лабораторных условиях и то лишь с большим трудом. Таким образом, чрезвычайно быстро сформировавшееся следование за объектом с определенными отличительными признаками в дальнейшем уже существенно не меняется. Лишь к концу действия запечатления (когда выводок распадается) ослабевает первоначальная интенсивность этой реакции, образовавшаяся без внешнего подкрепления и не нуждающаяся в таковом для своего сохранения. Именно такую характеристику давал процессам запечатления в свое время Лоренц.
Половое запечатление
По-иному проявляется запечатление в сфере размножения. Здесь у многих животных наблюдается так называемое половое запечатление, которое обеспечивает будущее общение с половым партнером. Главная особенность полового запечатления заключается в том, что окончательный результат проявляется лишь с большой отсрочкой, ибо животное учится распознавать отличительные признаки будущего полового партнера еще на раннем этапе постнатального развития. При этом запечатление типичных признаков полового партнера должно совершиться у детеныша в таком виде, в каком они предстанут перед ним уже во взрослом, половозрелом состоянии.
В основном половое запечатление наблюдается у самцов, причем запечатляются у них отличительные признаки их матерей в качестве «образцов» самок своего вида. Следовательно, на врожденное узнавание общих видотипичных признаков здесь путем облигатного научения (полового запечатления) накладывается распознавание видотипичных женских признаков. Так еще в раннем возрасте происходит объективация будущего полового поведения.
В настоящее время факты полового запечатления установлены уже у различных позвоночных, но в первую очередь у птиц. Так, немецким этологом Ф.Шутцем было, в частности, показано, что у самцов диких уток сенсибильный период полового запечатления простирается от 10-го до 100-го дня с момента вылупления, но оптимальный срок ограничен 10–40 днями (в естественных условиях утиная семья распадается на 8–10-й неделе после появления утят). По другим данным этот срок может быть и несколько более длительным. Если на протяжении этого срока птенец мужского пола видел только самку другого вида, он по достижении половозрелости будет «ухаживать» только за самками этого вида, оставляя без внимания уток своего собственного вида. Из гусей, по данным Шутца, половое запечатление лучше всего проявляется у серого, а также и белолобого гуся.
Исследованиями шведского этолога Л. Б. Фэлта было установлено, что и у птенцов диких уток женского пола возможно половое запечатление, если их изолировать от селезней на протяжении первых восьми недель после вылупления. Если им при этом дать возможность видеть самцов-альбиносов или даже людей, то впоследствии нормальное половое поведение уток будет направлено на них, но не на селезней в их обычном брачном наряде. Половое запечатление на человека у представителей этого отряда (гусиных) в свое время было описано и Лоренцем. Известен аналогичный случай и в отношении самца рыжей цапли, выросшего с 10-дневного возраста в домашних условиях и проявившего все формы брачного поведения по отношению к человеку, вырастившему его.
Половое запечатление на альбиносов (у птенцов мужского пола) удалось экспериментально доказать и у зебровой амадины (вьюрка), т.е. незрелорождающейся птицы: птенцы, выращенные родителями-альбиносами, предпочитали впоследствии спариваться с самками-альбиносами, а не с нормально окрашенными особями. Аналогичные результаты были получены и в экспериментах над голубями, когда птенцы белой окраски первоначально выращивались вместе с темноокрашенными взрослыми и наоборот. После изоляции, длящейся с 40-дневного возраста до половозрелости, большинство самцов выбирало самок того цвета, который был свойствен особям, с которыми они в первые недели своей жизни выращивались.
Еще более четкие результаты дают опыты с заменой кладок, при которых птенцы с самого рождения выращиваются приемными родителями. При такой постановке эксперимента у голубей удавалось получить половое запечатление даже в отношении чужого вида. Интересно, что эффект этого запечатления постепенно ослабевает, хотя в какой-то мере еще обнаруживается даже у 5-летних голубей. Таким же способом была установлена возможность полового запечатления на чужой вид у двух вьюрков, легко размножающихся при клеточном содержании (Lonchura striata, Taeniopygia castanotis). И у этих птиц половое запечатление в полной мере проявляется у самцов, которые игнорируют в половозрелом состоянии самок собственного вида, но у самок половое поведение в значительно большей мере определяется врожденными механизмами (К.Иммельманн). Е.В.Лукина наблюдала нечто подобное у канарейки и чижа.
Половое запечатление вообще является весьма сложным процессом и по-разному проявляется у разных видов птиц. Например, оно играет, очевидно, меньшую роль у куриных, как это вытекает из экспериментов П. Гитона, показавшего, что условия выращивания не сказываются на последующем выборе петухами самок. Узнавание отличительных признаков противоположного пола является у этих птиц, очевидно, вполне врожденным.
У млекопитающих половое запечатление изучено главным образом у копытных и грызунов. Имеются, например, указания на то, что скрещивание между ламами викуньей и альпакой возможно лишь в том случае, если новорожденному представителю первого вида дают в качестве кормилицы самку альпака. Немецкие этологи Д. и Х. Мюллер-Шварце показали, что у чернохвостого оленя в половом запечатлении главную роль играют запахи. Группой итальянских исследователей проводились опыты, в которых мыши-родители опрыскивались пахучими веществами. В результате их детеныши, будучи половозрелыми, оказались не в состоянии различать пол сверстников, т.е. не могли найти полового партнера. Аналогичные результаты были получены и у крыс. Опыты с применением «приемных родителей» производились и с морскими свинками. Эффект полового запечатления на чужой вид был выявлен у самцов, отделенных в течение первой недели жизни от своих матерей.
Интересно отметить, что половое запечатление может иметь место и у взрослых особей. Доказано это для некоторых рыб, например меченосца (Xiphohorus maculatus) – известной аквариумной рыбки из отряда карпозубых. Самцы этого вида, как показали шведские этологи А. Фернё и С.Сьёландер, предпочитают самок той окраски, которую им приходилось видеть в течение двух месяцев после наступления половозрелости, но не раньше, как это имеет место в описанных выше случаях полового запечатления.
Подводя итог обзору процессов запечатления, важно подчеркнуть, что общим и наиболее существенным для всех форм запечатления является быстрая постнатальная достройка врожденного поведения, точнее, врожденных пусковых механизмов путем их дополнения индивидуально приобретаемыми компонентами. В результате инстинктивное поведение конкретизируется на определенных, индивидуально опознаваемых объектах, чем и обеспечивается эффективность выполнения инстинктивных действий.
Раннее факультативное научение
Как самостоятельная категория приобретения индивидуального опыта факультативное научение играет в раннем онтогенезе еще значительно меньшую роль, чем на последующих этапах онтогенеза. Это объясняется тем, что оно первоначально лишь дополняет облигатное научение.
Экспериментально факультативный компонент раннего научения можно выявить, воздействуя на детенышей искусственными стимулами, которые или совершенно не встречаются в нормальной среде обитания данного вида, или хотя и могут встречаться, не обладают сколько-нибудь существенной биологической валентностью и не обусловливают определенную инстинктивную реакцию. Кроме того, этот компонент может быть выявлен путем обучения животных действиям, не представленным в их видотипичном поведении.
В качестве примера можно привести эксперимент, в котором крысят в возрасте 5, 7, 9 и 11 дней обучали с помощью электрического раздражения преодолевать коридор, причем ток выключался только при достижении крысятами стенки в конце коридора. При всех прочих оборонительных реакциях (поворотах, попытках забраться на стенку, опрокидывании на бок) действие болевого раздражителя продолжалось. У всех подопытных животных по мере тренировки число таких реакций уменьшалось, но только 7– 11 -дневные крысята научились все быстрее находить правильное направление к стенке в конце коридора и тем самым скорее избавляться от действия тока. Это говорит о том, что до 5-дневного возраста факультативное научение проявляется в еще весьма ограниченных рамках и неспособно помочь животному в выборе адекватного направления действия. Еще позже у крысят появляются факультативные реакции на положительные раздражители. Так, например, приблизительно только с 20-дневного возраста их можно научить нажимать на рычаг за пищевое вознаграждение. Однако, по данным ряда исследователей, оптимальный период раннего постнатального научения начинается у них в месячном возрасте.
У хищных млекопитающих отмечаются аналогичные сроки формирования первых положительных навыков. У щенков было установлено, что прогрессивное развитие способности к формированию навыков в раннем постнатальном онтогенезе непосредственно зависит от развития кратковременной памяти, что в свою очередь связано с развитием коркового торможения (Р.Г.Лозовская и Г.А.Образцова).
У других незрелорождающихся млекопитающих, которые появляются, однако, на свет с уже вполне функционирующими дистантными рецепторами, факультативное научение начинается значительно раньше. Так, по данным советской исследовательницы Н.А.Тих, много лет изучающей психику обезьян в сравнительно-психологическом аспекте, у низших обезьян в условиях искусственного выращивания уже на 3–4-й день после рождения появляются специфические реакции на вид рожка, из которого кормят детеныша, в частности своеобразные движения рук для его обхватывания (после 40–50 кормлений). Экспериментальный условный рефлекс на звук Тих удалось выработать у детеныша гамадрила в возрасте 29 дней.
Необходимо, конечно, учесть, что первые условные рефлексы на тактильные, термические или проприоцептивные раздражители образуются и у других млекопитающих в первые же дни после их рождения. Не исключено, следовательно, что в сфере действия таких раздражителей с самого начала постнатального развития может иметь место не только облигатное, но и факультативное научение даже у животных, рождающихся в крайне незрелом состоянии. Само собой разумеется, что именно так обстоит дело у всех зрелорождающихся животных.
Что касается низших позвоночных, то, как показали некоторые исследования, в частности В.А.Трошихина, экспериментальная выработка стойких условных рефлексов на искусственные раздражители удается уже у мальков рыб. Так, для образования условного пищевого двигательного рефлекса на вспыхивание электрической лампочки у малька карпа достаточно в среднем 4–6 сочетаний, причем положительная условная реакция остается стойкой в течение 100 повторений светового раздражителя без пищевого подкрепления.
Взаимоотношения между компонентами раннего постнатального развития поведения
Все компоненты раннего постнатального развития поведения переплетаются друг с другом, в результате чего это развитие приобретает очень сложный характер. Проанализируем эти связи и взаимозависимости на приведенном выше примере поведения вылупившихся гоголят.
В поведении этих птенцов прежде всего обращает на себя внимание врожденная (инстинктивная) двигательная реакция – подскакивание птенцов к выходу из дупла в ответ на призывные крики матери, а также врожденное узнавание светлого отверстия как цели прыжка. Врожденный характер подскакивания подтверждается работами норвежского исследователя Т.Бьерке, установившего, что для его появления необходим определенный уровень физиологической «зрелости» птенца. Но едва ли имеет здесь место также всецело врожденное узнавание призывного крика, так как еще до вылупления птенцов между ними и материнской особью, вероятно, устанавливается уже знакомый нам взаимный акустический контакт, как это доказано для других зрелорождающихся птиц. Следовательно, нельзя исключить возможность своего рода акустически-кинестетического эмбрионального запечатления голоса матери, т.е. пренатального облигатного научения, как его теперь можно назвать. С другой стороны, совершенно очевидно, что птенцы не могли в период эмбриогенеза тренироваться в целенаправленных прыжках.
Далее существенно, что гоголята реагируют на звуки матери еще до того, как увидят ее: только высунувшись из дупла, они впервые видят ее летящей перед собой. Также врожденная реакция спрыгивания на землю непосредственно не связана с ее присутствием (хотя, возможно, и стимулируется ее видом и голосом), так как подобным же образом ведут себя и гоголята, выращенные в инкубаторе, когда они оказываются на светлом месте (наблюдения Фабри).
Завершается этот первый постэмбриональный этап собственно реакцией следования утят за удаляющейся к водоему матерью (и друг за другом). Эта реакция наступает в ответ на вид двигающейся (в данном случае ходящей) птицы в сочетании с издаваемыми ею звуками, что четко доказано экспериментами, проведенными многочисленными исследователями не только на утятах, но и на цыплятах и птенцах других зрелорождающихся птиц.
Выше уже отмечалось, что конкретное значение запечатления типа реакции следования состоит в индивидуальном узнавании особенно важного компонента среды – родительской особи. Ясно, что это не может осуществляться только врожденным образом, так как распознаваться и запоминаться должны в данном случае не видовые, а именно индивидуальные признаки, т.е. частные отклонения от видотипичной нормы. Объектами врожденного узнавания могут быть лишь общие всем представителям данного вида признаки, а между тем детеныш должен быть в состоянии отличить своего родителя как раз от других особей того же вида. Этому можно только научиться, на это и направлено облигатное научение.
В некоторых случаях, правда, видотипичные признаки, хотя бы частично, тоже фиксируются в памяти детеныша путем запечатления. Особенно, как мы видели, это проявляется при половом запечатлении. В этом состоит одно из важнейших отличий полового запечатления.
Важно отметить, что процессы раннего облигатного научения, прежде всего запечатления, всегда сочетаются с врожденным узнаванием. При этом врожденное узнавание направлено на общие существенные признаки объектов видотипичных действий, путем же облигатного научения эти действия конкретизируются на частном объекте. Так, при реакции следования восприятие детенышем самой подвижности родительской особи (как и любого другого впервые увиденного объекта) является врожденным. Это обусловлено тем, что подвижность представляет собой в данном случае существенный признак объекта запечатления. Индивидуальные же отличительные признаки родительской особи (т.е. объекта видотипичного действия) заучиваются путем запечатления. Следовательно, слияние врожденного поведения (врожденного узнавания) с облигатным научением служит основой дальнейшего развития видотипичного поведения.
Вместе с тем в запечатлении содержатся элементы и факультативного научения. Так, например, птенцы одного и того же выводка могут научиться узнавать родительскую особь по разным ее признакам, и при этом не имеет значения, по каким конкретно признакам ориентируется тот или иной птенец. Облигатным, обязательным здесь является то, что каждый птенец должен вообще научиться узнавать родителя по каким-либо его индивидуальным признакам.
Сочетание врожденных и приобретаемых компонентов сказывается здесь еще и в другом. Как показали эксперименты, проведенные Фабри на цыплятах, первоначальный эффект врожденного узнавания при реакции следования недолговечен и без постоянного упражнения довольно быстро угасает. Однако этот эффект, т.е. следование, можно легко восстановить путем факультативного научения, направленного на объект запечатления. В упомянутых экспериментах объектом запечатления служил для одних цыплят синий шар, для других – желтый цилиндр. После первых же предъявлений цыплята стали следовать за этими объектами не менее интенсивно и продолжительно, чем в обычных условиях за курицей. Но даже наиболее активные подопытные цыплята переставали следовать за объектом запечатления уже через несколько дней, если прекращались ежедневные тренировки. В обычных же условиях, как известно, эта реакция сохраняется в течение ряда недель. Однако и в искусственных условиях эксперимента цыплята тем дольше продолжали следовать за объектом запечатления, чем больше было всего пробежек, т.е. длительность сохранения реакции следования оказалась прямо пропорциональной интенсивности локомоторной активности (частоте пробежек).
В дальнейшем, уже после того, как реакция следования у цыплят совсем исчезла, у них в специальной установке были выработаны навыки следования за бывшим объектом следования – синим шаром или соответственно желтым цилиндром. Следование за этим объектом подкреплялось пищей. Другими словами, следование было заново выработано у цыплят, но путем факультативного научения.
Результаты этой части эксперимента показали, что у цыплят, у которых раньше была сформирована реакция следования путем запечатления, этот навык образовался намного быстрее, чем у цыплят, которые до этого вообще не подвергались запечатлению, т.е. не имели возможности следовать за каким-либо подвижным объектом или же следовали за объектом запечатления другой группы подопытных животных (для цыплят, запечатленных на синий шар, это был желтый цилиндр и наоборот, – рис. 21).
Из этих данных, казалось бы, следует, что врожденное действие (само следование), протекание которого не зависит от побочных внешних факторов и сроки проявления которого генетически фиксированы, угасает без упражнения так же, как это происходит с навыками, а с другой стороны, навык, факультативное научение, может вновь «оживить» эту инстинктивную реакцию. Объясняется это, очевидно, тем, что существует не одна, а две генетически различные, но функционально тождественные формы следования – истинно врожденная и индивидуально приобретаемая, причем вторая форма образуется путем чисто факультативного научения (ведь курица подкрепляет следование своих цыплят кормом и обогревом). Очевидно также, что первая форма со временем все больше перерастает во вторую, иными словами, реакция следования претерпевает качественную перестройку, теряя свою первоначальную врожденную сущность. Одновременно врожденное узнавание переходит в благоприобретаемое узнавание, основанное на индивидуальном опыте.
Рис. 21. Формирование следования за подвижным объектом путем запечатления и факультативного научения (опыты Фабри). А – экспериментальная группа цыплят, Б – контрольная группа цыплят. 1 – выработка и тренировка реакции следования за синим шаром путем запечатления (1–4-е сутки после вылупления);
2 – первый контрольный опыт; цыплята следуют за объектом запечатления (синим шаром), но не за другим подвижным объектом (желтым цилиндром) (5-е сутки); 3 – второй контрольный опыт (26-е сутки), проведенный после трехнедельного перерыва, во время которого цыплята находились в обычных условиях, но не имели возможности видеть шар и цилиндр. В результате цыплята не только не следуют за этими движущимися объектами, но даже пугаются их. Так же реагируют на движущиеся шар и цилиндр контрольные (не подвергшиеся запечатлению) цыплята; 4 – выработка навыка следования за двигающимися шаром или желтым цилиндром при одновременном их предъявлении (27–31-е сутки) путем факультативного научения. Столбики с цифрами показывают среднее число сочетаний, необходимых для формирования устойчивого следования
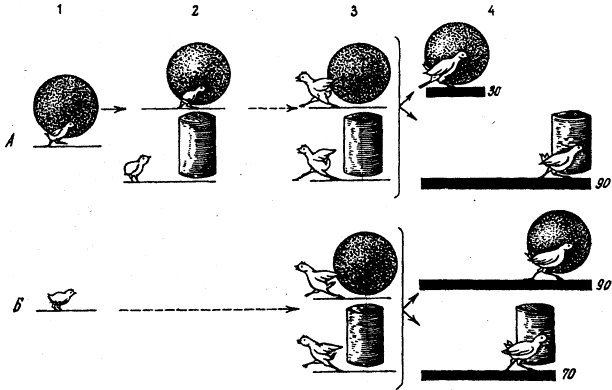
Все это указывает на значение раннего факультативного (а не только облигатного) научения для полноценного проявления инстинктивных элементов поведения. Иными словами, уже на первых этапах постнатального развития врожденное поведение нуждается в участии научения в обеих его формах, которые при этом сливаются с ним. Этим прежде всего и характеризуется сущность всего поведения животных на ранних этапах постнатального онтогенеза.
Раннее формирование общения
Раннее формирование общения у птиц
Ознакомившись в общих чертах с основными компонентами раннего постнатального развития поведения и их взаимоотношениями, рассмотрим теперь специально, как формируется общение между детенышами и родителями и другими особями. Правда, коммуникативные компоненты поведения уже неоднократно встречались нами при ознакомлении с разными аспектами онтогенеза поведения, особенно в процессах запечатления, одним из определяющих факторов которого и является общение.
У птиц, как было уже показано (в частности, в предыдущей главе на кайрах), установление акустических контактов между родительской особью и ее потомством начинается уже на эмбриональной стадии развития птенцов путем пренатального облигатного научения (эмбрионального запечатления). У кайр, например, этот процесс происходит в форме установления акустико-кинестетических связей. В результате этого запечатления птенец подбегает к источнику запечатленного звука. Эта реакция обнаруживается даже у только что вылупившегося птенца: услышав запечатленный звук, он подбегает к родительской особи (в эксперименте к замещающему ее громкоговорителю), издает ответный крик и залезает под нее (или под аппарат). Если же из громкоговорителя после этого раздается другой звук (чужой зов), то птенец немедленно выскакивает из-под него и прячется в укрытии (опыты Чанц). Таким образом, коммуникативный компонент врожденного пускового механизма достраивается в данном случае в ходе эмбрионального запечатления.
Вместе с тем и у родительской особи происходит запечатление индивидуальных особенностей голоса еще не вылупившегося птенца, что обеспечивает индивидуальное опознавание последнего уже к моменту вылупления. Подобное «обратное запечатление» было установлено и у других птиц, а в постнатальном периоде также у млекопитающих и ряда рыб.
Укажем еще на работы Готтлиба, который показал на утятах, что если эмбрион не подвергается в последние дни инкубации соответствующим акустическим воздействиям, то установление звуковых контактов между вылупившимся птенцом и материнской особью будет значительно затруднено. Изучая затем реакцию следования у нескольких видов уток (крякв, пекинских уток, мандаринок), Готтлиб установил, что наиболее активно утята следовали за макетом (чучелом) при видотипичной визуально-акустической стимуляции, хуже – при одной лишь акустической, т.е. без чучела, и еще хуже при одной оптической, т.е. при неозвученном чучеле. Вместе с тем реакция следования значительно ослабевала, когда видотипичные визуальные или акустические компоненты (позывные звуки и внешний вид чучела) подменялись невидотипичными (т.е. принадлежащими другому виду уток). Канадский ученый Р.М.Эванс в опытах над чайками показал, что искажения в реакциях вылупившихся птенцов (предпочитание криков другого вида) могут обусловливаться отсутствием соответствующего эмбрионального опыта, а именно когда эмбрион лишен возможности слышать крики птиц своего вида.
Итак, как правило, вылупившиеся птенцы реагируют четко избирательно на видотипичные крики, т.е. предпочитают ориентироваться по «натуральным» акустическим раздражителям с большой биологической валентностью. Эти реакции основаны на врожденном узнавании (и предпочтении) видотипичных компонентов коммуникации, которое дополняется и уточняется облигатным выучиванием индивидуальных компонентов общения в процессе пре- и особенно постнатального запечатления. Следовательно, при постнатальном формировании акустического общения между птенцами и их родителями возможность индивидуального (не врожденного!) опознавания особей обеспечивается ранним облигатным научением. Принадлежность же последних к одному и тому же виду устанавливается птенцами на основе врожденного узнавания.
На основе данных, полученных рядом ученых, сейчас можно, очевидно, считать установленным, что и другие функции пения птиц формируются в большой степени благодаря постнатальному облигатному научению. Например, у овсянки песня молодого самца приобретает сексуальную, а затем и территориальную функцию только после того, как он научается петь «полную» песню, т.е. модифицировать ювенильную «подпесню», придать ей специфическую структуру и видоизменить ритм.
Большую роль играет при этом подражание, на что, в частности, еще в середине 50-х годов указывал Торп, отмечая, что птицы учатся непосредственно по звуковым сигналам других птиц.
Выше уже рассматривались явления подражания и говорилось о необходимости выделить как отдельные категории облигатное и факультативное имитационное научение. Теперь должно быть ясно, что облигатное подражание, будучи особым проявлением облигатного научения, характерно прежде всего для раннего постнатального периода онтогенеза. В области звукового общения это можно хорошо проиллюстрировать примерами из многочисленных полевых экспериментов, осуществленных советскими орнитологами К.А. и К.К.Вилкс путем перемещения яиц и птенцов в гнезда других видов (еще раньше подобные эксперименты проводились Промптовым). Изучив поведение приблизительно 4000 пересаженных воробьиных птиц (преимущественно мухоловок-пеструшек), эти исследователи установили, что птенцы в исключительно широком диапазоне перенимают песни «приемных родителей»: более 80% самцов, выросших в гнездах других видов, имитировали песню вида, к которому принадлежала особь, воспитавшая птенца (горихвостки, пеночки-трещотки, большой синицы). Вместе с тем при имитации чужой песни воспроизводились и элементы песни собственного вида. Понятно, что в нормальных условиях акустическое поведение птенца формируется путем облигатного имитационного научения звукам, издаваемым кормящей птицей того же вида.
Примечательно, что перенимание чужих звуков происходит в относительно короткий период начиная с вылета птенца из гнезда. Это доказывается тем, что чужие звуки имитировали и птицы, пересаженные незадолго перед вылетом, но если, наоборот, птенец воспитывался с самого начала в чужом гнезде, но перед вылетом пересаживался обратно к своему виду, чужое воспитание не сказывалось на акустических реакциях такой птицы и ее пение оставалось видотипичным. Таким образом, здесь определенно можно говорить о наличии сенсибильного периода, что опять-таки доказывает, что мы имеем в этом случае дело с облигатным научением, точнее, облигатным имитационным научением.
Раннее формирование общения млекопитающих
У млекопитающих взаимное запечатление индивидуальных отличительных признаков родителей и детенышей и установление контактов между ними происходят в разные сроки после появления детеныша на свет – в зависимости от степени зрелорождения, но чаще всего в первые часы жизни новорожденного. Детеныш одногорбого верблюда, например, издает первые звуки еще во время самих родов, а уже час спустя он в состоянии воспроизвести почти все звуки, свойственные его виду, поэтому уже с момента рождения начинается интенсивный акустический контакт между ним и его матерью.
Американским исследователем онтогенеза поведения животных Дж. П. Скоттом и рядом других ученых было доказано, что у собак и других видов псовых существует сенсибильный период, в течение которого только и возможно такое установление контактов с материнской особью и остальными щенками данного выводка. Очевидно, при этом процесс запечатления имеет комплексный характер, поскольку включает в себя и половое запечатление. На это указывает и то обстоятельство, что общение между матерью и щенками, равно как между последними и другими представителями того же вида, может быть по форме идентично половому общению (М. В. Фокс). Так же, по свидетельству ряда ученых, обстоит дело и у обезьян.
Большое значение для формирования общения имеет положительная тигмотаксисная реакция детенышей, выражающаяся в активном установлении физического контакта с поверхностью матери и собратьев, в прижимании к ним. Об этом свидетельствуют, в частности, эксперименты, проведенные на детенышах обезьян Г. Харлоу и его сотрудниками, применявшими разного рода макеты материнских особей (проволочные, тряпочные, обогревамые, холодные и пр.). Было показано, что детеныши предпочитают запечатленный холодный макет даже обогреваемому. Сходная тигмотаксисная реакция, как показали английский этолог В. Слэкин и другие исследователи, присуща также цыплятам и другим птенцам.
Скотт пришел к выводу, что у млекопитающих в раннем постнатальном онтогенезе существуют сенсибильные периоды трех типов: для процессов научения, стимуляции физиологических процессов и формирования общения. Исследования многих ученых показали, что если в течение сенсибильного периода последнего типа детеныш не имеет возможности установить связи с особями своего вида, он впоследствии может оказаться совершенно неспособным к общению с себе подобными и во всяком случае будет испытывать большие затруднения в коммуникативном, а также воспроизводительном поведении. То, что формирование первичных коммуникативных связей совершается путем запечатления, подтверждается и тем, что оно, как доказано рядом экспериментаторов, не зависит от пищевого или иного внешнего подкрепления.
Не менее четко выражено у млекопитающих и «обратное запечатление» – запечатление индивидуальных признаков детенышей у родителей. Так, например, овцы, козы и другие копытные научаются распознавать эти признаки непосредственно после рождения их детенышей и после этого уже не подпускают к соскам чужих. Ведущее значение имеет здесь, вероятно, ольфакторное опознавание, т.е. запоминание индивидуального запаха потомства, хотя и оптические признаки могут играть существенную роль. У детеныша, как показали в эксперименте с чернохвостыми оленями Д. и Х.Мюллер-Шварце, также происходит запечатление индивидуального запаха матери или, в эксперименте, заменяющей модели, причем в отношении последней вполне осуществляются акты общения, проявляемые в норме по отношению к подлинной матери.
Формирование общения у низших позвоночных
Отмеченные закономерности не имеют конечно, значения всеобщих правил для всех позвоночных, а тем более беспозвоночных. Даже среди птиц и млекопитающих обнаруживаются многочисленные отклонения и исключения, обусловленные экологическими факторами, особенностями образа жизни. С другой стороны, эти закономерности проявляются и у многих рыб. Так, например, мальки цихлид также научаются опознавать своих родителей путем запечатления и неотступно следуют за ними, но только тогда, когда те находятся в брачном наряде. Таким образом, облигатное научение здесь более крепко связано с врожденным узнаванием и более ограничено непосредственным действием ключевых раздражителей (в данном случае светло-темные контрасты, зеленые и синие цвета, движение), чем это наблюдается у высших позвоночных. Доминирующую роль здесь играют врожденные пусковые механизмы, индивидуальное же распознавание играет второстепенную роль. Средствами общения являются при этом сами движения, выполняемые родителями и мальками и имеющие генетически фиксированное сигнальное значение.
Что же касается «обратного запечатления», то оно у цихлид не связано с индивидуальным опознаванием потомства родительскими особями. Последние узнают мальков лишь как таковых, но неспособны отличить свой выводок от чужого, как это показали опыты с перекрестным воспитанием молоди путем замены родителей в семейных группах. При этом более крупная молодь поедается. Если подрастающих мальков повторно заменить более молодыми (т.е. более мелкими), то уход за потомством со стороны родителей можно растянуть на срок до 9 месяцев. На этой фазе преобладает оптическое общение, на личиночной – химическое (опыты Д.Наукса и Дж.Барлоу).
У низших позвоночных, в частности у рыб, присутствие особей того же вида не является столь необходимым для формирования форм общения, как у высших животных. В этом также проявляется более жесткое врожденное «программирование» раннего постнатального поведения рыб, чем у последних. Сходное положение наблюдается, очевидно, и у пресмыкающихся. Так, у ящериц-анолисов важный компонент общения – кивание головой с одновременным вытягиванием ног – наблюдается уже у новорожденных (через 5 минут после вылупления), причем и у изолированных животных, лишенных возможности видеть сородичей. Таким образом, здесь налицо подлинные врожденные двигательные координации, жестко регламентирующие коммуникативное поведение без индивидуального опознавания сородичей.
Познавательные аспекты раннего постнатального поведения
Исследовательское поведение
Постоянной составной частью любого поведенческого акта является исследовательское поведение, проявляющееся на разных онто- и филогенетических уровнях в весьма различных формах. Диапазон этих проявлений простирается от элементарных ориентировочных реакций до исследовательской деятельности высших млекопитающих.
В широком смысле ориентировочное, поисковое поведение является начальной фазой любого инстинктивного действия, и оно направлено на поиск тех агентов среды, которые являются объектами врожденного узнавания. Иначе говоря, в инстинктивных поведенческих актах ориентировочно-исследовательская активность служит поиску и обнаружению ключевых раздражителей и направляет на них деятельность животного. Об этом уже говорилось при изложении этологической концепции инстинктивного поведения.
Элементарные ориентировочные реакции проявляются очень рано даже у незрелорождающихся детенышей. Так, у щенят и лисят уже в первый-второй день жизни наблюдаются поисковые маятникообразные движения головой, которые прекращаются после нахождения соска матери. Сходные поисковые движения, повороты головы в стороны и тыканье концом морды в окружающие животное объекты производятся в первые же сутки жизни также котятами и многими другими детенышами хищных млекопитающих. Детеныши обезьян уже в первые дни жизни двигают ушными раковинами и т.д.
В дальнейшем появляются такие ориентировочные реакции, как обнюхивание, прислушивание, присматривание и т.п. У щенят и лисят такое обследование окружающего пространства начинается приблизительное конце второй недели жизни. У детенышей низших обезьян начиная с 14–16-го дня жизни появляются двигательные реакции в ответ на «индифферентные» звуковые и зрительные раздражители, которые потом (приблизительно в 1,5–2-месячном возрасте) вырастают в подлинные исследовательские действия: животное живо реагирует на изменения в окружающей среде (появление нового раздражителя, нового объекта) поворотами головы, особенно глаз, движениями ушных раковин, а также приближением к новому, незнакомому объекту. Так появляется «любопытство», столь характерное прежде всего для приматов, а также для других высших позвоночных.
Развитие исследовательского поведения в большой мере определяется условиями, в которых растет детеныш, особенно возможностями общения с матерью и другими сородичами. В экспериментах с длительным (до 30 недель) изолированным выращиванием детенышей макаков-резусов (опыты американских ученых Ч.М.Байзингера, Е.М.Брандта и Дж.Мичелля) было установлено, что изолированные детеныши больше рассматривали окружающие их предметы, человека, наблюдавшего за ними, и самих себя, чем нормально живущие со своими матерями. Первые также больше манипулировали предметами и больше трогали самих себя, вторые больше, быстрее и ловчее передвигались по клетке. Чехословацкие исследователи Б.Семигиновский, З.Халупка и И.Мысливечек, ставя опыты над крысятами, нашли, что возможность усиленного общения с сородичами в течение первых 6 недель жизни несколько тормозит развитие исследовательской активности, а обогащенная среда, наоборот, стимулирует его.
Уже на начальных стадиях постнатального онтогенеза наряду с реакциями на появление новых раздражителей имеет место активный поиск последних (маятникообразные движения головой у щенков), который особенно характерен для исследовательского поведения. Лежащие в основе этого поведения ориентировочные рефлексы отличаются, как показали исследования Е.Н.Соколова, значительной сложностью их физиологических механизмов. По Соколову, ориентировочные реакции возникают в результате несовпадения воспринимаемых новых сигналов с формирующимися в головном мозгу «нервными моделями стимула». При этом происходит оценка нового стимула, побуждающая животное к выполнению определенных действий или к их изменению. Наряду с этим ориентировочная деятельность играет также существенную роль в обеспечении необходимого уровня физиологической активности центральной нервной системы.
Нас же в данном случае особенно интересует познавательное значение ориентировочно-исследовательской деятельности, ее роль в приобретении новой информации о компонентах окружающей среды, в обогащении индивидуального опыта животного.
Облигатное научение и ориентация
Рассмотрим сначала некоторые процессы, связанные с начальной ориентацией у детеныша. У всех животных здесь первостепенное значение имеют таксисы, которые, как уже было показано, у высших животных дополняются и обогащаются элементами научения.
Это научение носит прежде всего характер облигатного научения, ибо, как только что отмечалось, оно входит как обязательный компонент в любой инстинктивный акт. Однако каждое животное вынуждено самостоятельно выучивать значимые лишь для него ориентиры, которые будут для разных особей того же вида весьма различными. Отличительные признаки этих ориентиров являются сами по себе случайными, несущественными, и только индивидуальное запоминание их в результате факультативного научения придает им опознавательное значение. Уже говорилось о том, что их выбор животным определяется в большой степени индивидуальными особенностями последнего и разные особи могут предпочитать разные компоненты среды в качестве ориентиров.
Таким образом, в ориентировочном поведении детеныша всегда присутствуют элементы облигатного и факультативного научения, однако соотношения между этими двумя компонентами, их удельный вес могут быть различными в зависимости от того, в какой функциональной сфере совершается ориентация. Когда детеныш учится распознавать «живые ориентиры», особенно внешние признаки своих сородичей, т.е. когда ориентация сочетается с общением, явно преобладают моменты облигатного научения. Это вполне понятно, ибо когда животное ориентируется по подаваемым другими животными оптическим, акустическим, ольфакторным и иным сигналам, которые входят в репертуар общения, то решающее значение имеют наследственно фиксированные, врожденные элементы поведения. При формировании коммуникативного поведения в онтогенезе первостепенное значение имеет поэтому срочная постнатальная достройка соответствующих врожденных пусковых механизмов, что, как мы уже знаем, является главной характерной чертой запечатления. Ограничимся здесь отсылкой и к другим аспектам и примерам облигатного научения с ориентационным значением (например, привыкание к «биологически нейтральным» раздражителям), о которых речь шла выше.
Раннее факультативное научение и ориентация
Уже в раннем ориентировочном поведении заметно сказываются индивидуальные особенности животного. В большой степени индивидуальные различия в поведении зависят от частоты и характера осуществленных с момента рождения сенсорных действий. Это достаточно четко проявляется при выращивании детенышей в условиях, когда им приходится постоянно видеть определенные фигуры. Как показали экспериментальные исследования, животные, выросшие в таком окружении, впоследствии легче ориентируются по таким фигурам. Правда, согласно экспериментам американского ученого Р.М.Осволта, существенное значение имеет при этом степень трудности различения таких фигур. В этих экспериментах крысята еще до прозрения помещались в клетки, на стенах которых попарно помещались фигуры: для одних это были горизонтальные и вертикальные полосы, для других – дуги, для третьих – треугольники и круги. Заранее было известно, что последнее сочетание является наиболее трудным для различения этими животными, полосы же – наиболее легким. В таком окружении крысята пребывали до 50-дневного возраста. Контрольные животные выращивались в клетках без фигур (с белыми поверхностями). Когда затем перед крысятами была поставлена задача – произвести выбор между знакомыми фигурами (одна из которых подкреплялась пищей, другая – нет), то оказалось, что ранний сенсорный опыт облегчал решение задачи только тогда, когда животным приходилось ориентироваться по трудноразличимым фигурам (треугольник и круг). Выращивание же с легче различимыми фигурами не давало такого эффекта: соответственные подопытные животные ориентировались по полосам и дугам не лучше контрольных. При этом Осволт отмечает, что влияние «преэкспозиции» стимулов явно приурочено к определенному раннему периоду, что указывает на участие в данном процессе наряду с факультативными также облигатных компонентов латентного научения.
Итак, ранняя факультативная неподкрепляемая визуальная тренировка способствует ориентации при дальнейшем подкрепляемом факультативном научении (заучивании ориентиров), особенно тогда, когда эта тренировка была сопряжена с трудностями и если имели место более сложные сенсорные действия.
В целом приведенные факты ранней пространственной ориентации показывают, как облигатное научение (благо приобретаемая ориентация по биологически валентным элементам среды) сочетается с факультативным научением, дополняется и конкретизируется им (запоминание особенностей местности путем сенсомоторной тренировки и заучивание конкретных ориентиров). При пространственной ориентации описанного типа на первый план выступает факультативный компонент. Иное соотношение наблюдается, как мы видели, в тех случаях, когда животное ориентируется не по признакам местности, а по признакам сородичей или других животных.
Раннее манипулирование
Исключительно большое значение имеет для приобретения и обогащения индивидуального опыта, как и для всей познавательной деятельности животного, манипулирование. Под манипулированием, манипуляционной активностью мы понимаем активное обращение с различными предметами при преимущественном участии передних, реже – задних конечностей, а также других эффекторов: челюстного аппарата, хобота (у слона), хватательного хвоста (у широконосых обезьян), щупалец (у головоногих моллюсков), клешней (у раков) и т.д. Манипулирование проявляется прежде всего в пищедобывательной и гнездостроительной активности животных. Вообще же, за исключением некоторых ориентировочных, сигнальных и защитных движений, при которых меняется лишь внешний вид животного, но само оно остается на месте, все движения, направленные на внешнюю среду, т.е. составляющие поведение, подразделяются на локомоторные (у большинства высших животных – опорно-локомоторные) и манипуляционные.
У высших животных, как еще будет показано, манипулирование выступает как ведущий фактор развития сенсомоторных функций. Это обусловлено тем, что именно в ходе манипулирования животное вступает в наиболее активный контакт с предметными компонентами среды и получает наилучшие возможности для ознакомления с ними, а также для многообразного воздействия на них.
Манипулирование появляется в различные сроки и проявляется по-разному у разных животных. Особенно велики эти различия у зрело- и незрелорождающихся видов. Но в каждом случае, выполняя разнообразные действия с различными предметами (биологически значимыми и «нейтральными»), детеныши получают комплексную информацию об объекте манипулирования, его свойствах, особенно о его физической структуре. Одновременно происходит развитие и совершенствование (путем тренировки) эффекторносенсорных систем животного. Все это делает манипуляционную активность высшей формой ориентировочно-исследовательской деятельности животных.
В полной мере манипулирование развертывается в игровом периоде онтогенеза. В рассматриваемом здесь периоде манипуляционная активность появляется первоначально лишь в самых простых формах, особенно у незрелорождающихся животных. Так, например, детеныши хищных млекопитающих в первые дни или даже недели, а именно до начала функционирования дистантных рецепторов, т.е. до открывания глаз и ушных проходов, в основном спят, а в промежутках преимущественно ползают в поисках соска и сосут.
По данным советского этолога Н.Н.Мешковой, первые движения детенышей носят характер простейшего манипулирования: детеныши трогают лапой родителей и своих собратьев, хватают их ртом в разных местах тела, нажимают на них мордочкой. Все эти действия еще весьма нечеткие и непродолжительные. У лисенка, по исследованиям того же автора, до 12-дневного возраста (до прозрения) манипуляционная (нелокомоторная) активность развивается следующим образом.
Первые манипуляционные движения обнаруживаются уже у новорожденного, но в течение первых двух часов после рождения они существуют лишь в двух формах: 1) прикасания к объекту передним концом головы (конкретно это проявляется в поиске соска) и 2) хватания объекта губами (захват соска и прилегающих участков кожи ртом). Передние конечности в этих действиях участия не принимают и отведены в стороны.
Затем в течение первых двух суток с момента рождения к этим двум формам добавляются еще пять, которые на этом этапе также связаны только с сосанием и сопутствующими ему действиями (расталкивание других детенышей при движении к соску, поиск соска, отгребание шерсти на животе матери, придавливание или ритмичное толкание лапами живота рядом с соском во время сосания). Эти манипуляции характеризуются прежде всего тем, что детеныш начинает действовать передними лапами и появляются боковые движения головы (в частности, при расталкивании собратьев). Передние лапы прикасаются к объекту (телу матери) и передвигаются по нему одновременно с головой или же самостоятельно, без ее участия.
Последнее имеет место в тех случаях, когда лисенок разгребает одними передними лапами шерсть на животе матери или придавливает его сразу обеими лапами.
Далее, вплоть до прозрения, положение существенно не меняется, появляется лишь еще одна форма манипулирования – захватывание объекта с поочередным придавливанием его обеими передними конечностями. Это имеет место тогда, когда лисенок сосет и при этом ритмично нажимает на живот рядом с соском то одной, то другой лапой.
Итак, к моменту открывания глаз лисенок обладает лишь восемью формами манипулирования, которые к тому же выполняются преимущественно головой. Передние конечности играют в основном лишь вспомогательную роль и способны лишь к самым элементарным самостоятельным движениям. Действия, выполняемые лишь одной передней конечностью, на этом этапе онтогенеза еще полностью отсутствуют. Все это свидетельствует о том, что манипуляционные функции головы, в частности челюстного аппарата, опережают в своем развитии формирование функциональной системы передних конечностей, которые на первых порах еще недостаточно сильны для самостоятельного манипулирования предметами (из восьми форм лишь одна выполняется только конечностями). Здесь, как и в дальнейшем ходе онтогенеза, отчетливо проявляются уже отмеченные раньше реципрокные отношения между этими двумя эффекторными системами.
Познавательное значение раннего манипулирования
Как видно из приведенного примера, активность незрелорождающегося детеныша млекопитающего направлена на этом, доигровом, периоде онтогенеза практически только на один объект – тело матери. Лишь попутно начинают появляться действия, направленные на собратьев, и то лишь для устранения помех на пути к соску. Подлинно активного контактирования между детенышами нет (устанавливается в основном пассивный контакт во время совместного отдыха и сна). Активные действия, направленные на другие, особенно «биологически нейтральные», объекты, еще отсутствуют полностью. Это обстоятельство резко сужает познавательное значение манипуляционной активности на раннем этапе онтогенеза, так как детеныш получает лишь ограниченную и однообразную информацию.
. Совершенно иная картина наблюдается, естественно, у зрелорождающихся животных, которым раннее развитие эффекторных и сенсорных способностей позволяет уже в первые дни после появления на свет вступать в многообразные отношения с компонентами среды. Этап раннего формирования манипуляционной активности и связанного с этим постепенного созревания познавательных способностей у этих животных выпадает. О биологическом значении этой специфической особенности зрелорождения уже говорилось в начале главы в связи со взглядами Л. А. Орбели.
Несмотря на первоначальную ограниченность познавательной функции манипуляционной активности у незрелорождающихся животных, мы имеем здесь дело с началом процесса, который в ходе онтогенеза приобретает все большее значение для психического развития особи (см. следующую главу). В этой связи интересно отметить, что зрелорождающиеся животные (птицы и млекопитающие) вообще сравнительно мало манипулируют, во всяком случае в ходе дальнейшего онтогенеза этот компонент их поведения, как и другие, сравнительно мало прогрессирует.
Особенно четко значение манипулирования выступает при сравнении его с запечатлением. И в этом и в другом случае мы имеем внешне не подкрепляемую деятельность, причем генетически фиксированную, врожденную (все животные одного вида манипулируют в целом одинаковым образом). И в том и в другом случае эта деятельность специально направлена на распознавание элементов окружающего мира, на различение знакомого и незнакомого (при запечатлении – «своя» мать и «не свои» особи), на формирование индивидуального опыта. В обоих случаях имеет место упражнение и совершенствование эффекторносенсорных способностей, «моторное» и «сенсорное» научение: животные учатся адекватно употреблять свои органы движения и чувств, причем научение это совершается комплексно.
Но существенная разница между запечатлением и манипулированием состоит в том, что запечатление строго фиксировано тем, что оно направлено лишь на объекты видотипичных, инстинктивных действий, в то время как манипулирование не ограничено никакими рамками: объектом манипулирования может быть любой объект вообще. Более того, особая роль манипулирования в развитии психики и заключается в том, что она лишь первоначально в начале постнатального онтогенеза, направлена исключительно на биологически высоко валентные объекты, т.е. объекты инстинктивных действий. Полностью же и в наиболее сложных формах манипуляционная активность развертывается тогда, когда предметная деятельность подрастающего животного распространяется и на «биологически нейтральные» объекты. Это происходит, когда молодое животное начинает играть. Именно тогда начинает сказываться ведущее познавательное значение манипуляционной активности, ее роль как важнейшего фактора «латентного научения», накопления индивидуального опыта «впрок».
Глава 4 РАЗВИТИЕ ПСИХИЧЕСКОЙ ДЕЯТЕЛЬНОСТИ В ЮВЕНИЛЬНОМ (ИГРОВОМ) ПЕРИОДЕ
Общая характеристика игры у животных
Как уже отмечалось (см. ч. 2, гл. 1), о ювенильном (или игровом) периоде развития поведения можно говорить только относительно детенышей высших животных, у которых развитие поведения совершается перед половым созреванием в форме игровой активности. Поэтому только о таких животных и пойдет здесь речь. У других животных – а их подавляющее большинство – индивидуальное развитие поведения ограничивается описанными в предыдущей, главе процессами созревания врожденных форм поведения, облигатного и факультативного научения, равно как элементарными формами исследовательского поведения.
Игры животных давно привлекают внимание исследователей, но тем не менее они до сих пор еще плохо изучены. Сформулированные в разное время взгляды по этому вопросу в основном можно объединить вокруг двух концепций, впервые выдвинутых еще в прошлом веке, с одной стороны, Г.Спенсером, а с другой – К.Гроосом.
В первом случае игровая активность животного рассматривается как расход некоей «избыточной энергии», как своего рода суррогат «естественного приложения энергии» в «настоящих действиях» (Спенсер). Этот взгляд, акцентирующий эмоциональные аспекты игры, получил новое воплощение в современных представлениях о «вакуумной активности», наглядным примером которой являются описанные Лоренцем «действия вхолостую», т.е. инстинктивные движения, выполняемые при отсутствии соответствующих ключевых раздражителей. Правда, Лоренц указал и на существенные различия между игрой и «вакуумной активностью».
Основной недостаток трактовки игры у животных как реализации «избыточной энергии» заключается, как отмечал С.Л. Рубинштейн, в отрыве этой формы активности от ее содержания, в ее неспособности объяснить конкретные функции игры в жизни животных. Вместе с тем этологическая концепция о «действиях вхолостую» проливает некоторый свет на возможные элементы эндогенной мотивации игрового поведения животных.
Во втором случае мы имеем дело с «чисто функциональной» трактовкой игровой деятельности как упражнения в особо важных сферах жизнедеятельности. Вслед за Гроосом игру рассматривал как «практику для взрослого поведения» и К.Ллойд-Морган. Он подчеркивал, что игра позволяет молодому животному без риска упражняться в жизненно важных действиях, ибо в этих условиях ошибки не влекут за собой пагубных последствий: в ходе игры возможно совершенствование наследственных форм поведения еще до того как недостатки поведения роковым образом «предстанут перед судом естественного отбора».
Ясно, что полноценная теория игр животных должна включить в себя синтез положительных моментов концепции обоих этих направлений. Тем не менее до настоящего времени одни исследователи решительно отрицают функциональное значение игр молодых животных для формирования взрослого поведения, другие, наоборот, видят в последнем все значение игр. При этом отрицательная оценка зачастую сопровождается ссылкой на возможность созревания взрослого поведения без упражнения в ювенильном возрасте.
Так, известный голландский зоопсихолог Ф.Бойтендайк, выступая против концепции Грооса, утверждал, что игра важна только непосредственно для играющего, приводя его в положительное эмоциональное состояние, но не для его будущего. Инстинктивные формы поведения, по Бойтендайку, созревают независимо от упражнений; там же, где наблюдается упражнение в каких-то действиях, это не игра. Критику концепции Бойтендайка дал Д.Б.Эльконин, указав, в частности, на то, что Бойтендайк недооценил ориентировочно-исследовательскую функцию игры.
Значение игры для формирования взрослого поведения животных отрицается также и некоторыми другими зоопсихологами. Ряд ученых оставляют вопрос об упражняющей функции игры открытым (например, П.Марлер и В.Гамильтон), другие видят в игре некую «пара-активность» (А.Броунли), неспецифическую «мнимую деятельность» (М.Мейер-Хольцапфель), «самоподкрепляющуюся активность» (Д.Морис), «образцы» взрослого поведения (К.Лойзос) и т.д. Мейер-Хольцапфель называет одним из критериев игрового поведения «пробование», основанное на любопытстве. Существенно также, что игра имеет место лишь тогда, когда не выполняются подлинные инстинктивные действия. Темброк также отстаивает «автономность» игровой активности и считает возможным дать лишь негативное определение игр как поведенческих актов, функцию которых, как он пишет, нельзя непосредственно уяснить ни из действия, ни из их результатов.
Все же Темброк допускает, что игра увеличивает альтернативы поведения особи по отношению к окружающему миру, а путем включения элементов научения, возможно, формируются новые системы поведения в моторной сфере. Вслед за Лоренцем Темброк сравнивает игру с «действиями вхолостую». Игра тоже является «незавершенной», но указанные действия определяются мощной внутренней мотивацией и выполняются в жестких, невариабельных формах, в то время как игра во многих случаях зависит от внешних факторов, в частности от объектов игры, и выступает в весьма лабильных формах. Вместе с тем и для игрового поведения существуют поисковые (подготовительные) фазы и собственные ключевые раздражители, как это имеет место при типичном инстинктивном поведении. Игра часто направлена на «биологически нейтральные объекты», которые в остальное время не привлекают внимания животного. В отличие от сходных инстинктивных действий (например, борьбы или ловли добычи) игровые действия выполняются многократно и неутомимо. В итоге Темброк считает игры своеобразными инстинктивными действиями с самостоятельными мотивационными механизмами.
Иного мнения придерживается швейцарский ученый Г.Хедигер. Подчеркивая факультативный характер игры, он указывает на то, что в отличие от всех инстинктивных действий для выполнения игровых действий не существует специальных эффекторных образований, не существует никаких особых «органов игры». При этом он ссылается и на работы другого швейцарского ученого-физиолога В.Р.Хесса, который установил, что в мозгу кошки нет «центра игры», в то время как для всех инстинктивных действий такие центры можно обнаружить путем введения микроэлектродов.
Опираясь на исследования, проведенные совместно со своими сотрудниками, А.Д.Слоним высказывает мнение, что в определенные сроки постнатального развития инстинктивные реакции вызываются подпороговыми внешними раздражениями или даже только внутренними стимулами, возникающими в самом нервно-мышечном приборе. Возникшая в последнем случае «спонтанная» деятельность и проявляется в игровой активности. Хотя эта активность и не зависит от внешней среды, она может усиливаться условными рефлексами или внешними воздействиями (например, температурными).
Возвращаясь к вопросу о функциональном значении игры, необходимо отметить, что в настоящее время большинство исследователей все же считают, что игра служит подготовкой к взрослой жизни и накоплению соответствующего опыта путем упражнения, причем как в сенсорной, так и в моторной сфере. В последнем случае информация, поступающая реафферентно непосредственно от эффекторных систем, обеспечивает их «налаживание», выработку оптимального «режима действий».
В этой связи уместно упомянуть о предположении, высказанном Элькониным, что игра препятствует чрезмерно ранней фиксации инстинктивных форм деятельности и развивает все необходимые для ориентации в сложных и изменчивых условиях афферентно-двигательные системы. Как ювенильное упражнение рассматривает игру также Торп. По Торпу, игра служит для приобретения животными навыков и для ознакомления с окружающим миром. Особое значение Торп придает при этом манипулированию предметами.
Что касается прямых экспериментальных доказательств значения игры для формирования взрослого поведения, то такие доказательства были получены рядом исследователей. Еще в 20-х годах было, например, установлено, что сексуальные игры молодых шимпанзе являются необходимым условием способности к спариванию у взрослых особей (исследования Г. Бингхэма). В дальнейшем этот факт получил многократное подтверждение. ФА.Бич, а также известный исследователь поведения обезьян Г.В.Ниссен высказали даже утверждение, что у обезьян половое поведение зависит преимущественно от научения, но не от инстинктивных начал. Аналогичным образом совместные игры готовят молодых обезьян к будущей стадной жизни (исследования Г.Харлоу, С.Дж.Суоми и др.).
Экспериментально удалось доказать, что и самцы норок научаются выполнять действия, относящиеся к репродукционному поведению путем сексуальных игр. Особенно это относится к ухаживанию и подготовке к спариванию. Само спаривание формируется у них из вполне врожденных движений и игрового опыта, накопленного молодыми самцами в ходе общения с половозрелыми самками.
Большой интерес представляют данные, полученные Ниссеном совместно с К.Л.Чау и Дж.Семмесом. Эти экспериментаторы лишили детеныша шимпанзе возможности играть с предметами, не ограничивая при этом движения рук, в частности кистей и пальцев. Впоследствии возможности употребления рук, а также координация их движений оказались весьма несовершенными. Обезьяна резко отстала от своих нормальных сверстников по способности к хватанию и ощупыванию, не была способна локализовать тактильные раздражения поверхности тела с помощью руки (или она это делала крайне неточно), и вообще все движения рук были чрезвычайно неуклюжи. Характерно, что в отличие от других обезьян она не умела цепляться за ухаживающего за ней служителя, не протягивала к нему руки. Полностью отсутствовало даже столь характерное для обезьян обыскивание – важная форма их общения.
Многообразие толкований игр молодых животных обусловлено в большой степени тем, что игровая активность животных представляет собой сложный комплекс весьма разнообразных поведенческих актов, в своей совокупности составляющих содержание поведения молодого животного на этапе онтогенеза, непосредственно предшествующем половой зрелости. Поэтому Фабри предложил концепцию, согласно которой игра является по своей сущности развивающейся деятельностью, охватывающей большинство функциональных сфер. При таком понимании игры как развивающейся деятельности достигается синтетический подход к проблеме игровой активности животных, объединяющий все отмеченные выше моменты, и вместе с тем становится очевидным, что игровая активность наполняет основное содержание процесса развития поведения в ювенильном периоде. Игра представляются не какой-то особой категорией поведения, а совокупностью специфически ювенильных проявлений обычных форм поведения. Иными словами, игра является ювенильной (можно сказать, и «преадультной», т.е. «перед взрослым состоянием») фазой развития поведения в онтогенезе.
По этой причине невозможно согласиться с упомянутыми выше взглядами на игру как на «пара-активность», «мнимую деятельность» или «образец» взрослого поведения. Такие толкования нельзя оправдать и ссылками на несовершенство, неполноту или неполноценность игровых действий, на отсутствие соответственного биологического эффекта, ибо все это как раз и указывает на то, что мы имеем здесь дело с процессами развития поведенческих актов или, что то же самое, с действиями, которые еще не достигли своего полного развития. Игра – это не «образец» взрослого поведения, а само это поведение в процессе своего становления.
Сказанное, разумеется, не означает, что формирование поведения взрослого животного совершается только путем игры или что игра идет на смену другим, более ранним компонентам онтогенеза поведения. Наоборот, и у высших животных, детеныши которых активно играют, в ювенильном периоде сохраняются и продолжают действовать описанные в предыдущей главе факторы раннего онтогенеза, но в ювенильном периоде они часто сливаются – обычно в трансформированном виде – с игровым поведением. Одновременно, конечно, совершенствуется поведение детенышей и в «неигровых» ситуациях, как это имело место на доигровом этапе онтогенеза.
Важно также подчеркнуть, что, как будет еще показано, в ходе игры развиваются и совершенствуются не целиком взрослые поведенческие акты, а составляющие их сенсомоторные компоненты. Вообще же игровая активность, осуществляясь на врожденной, инстинктивной основе, сама служит развитию и обогащению инстинктивных компонентов поведения и содержит элементы как облигатного, так и факультативного научения. Соотношение этих компонентов может быть различным в разных конкретных случаях. Но в целом можно сказать, что в игровой активности завершается длительный и чрезвычайно сложный процесс формирования элементов поведения, берущий свое начало от эмбриональных координации и ведущий через постнатальное созревание врожденных двигательных координации и накопление раннего опыта к формированию и совершенствованию двигательных координации высшего уровня. Этот последний этап развития двигательной активности и представлен игрой.
Наряду с этим игра выполняет весьма важную познавательную роль, особенно благодаря присущим ей компонентам факультативного научения и исследовательского поведения. Эта функция игры выражается в накоплении обширного индивидуального опыта, причем в ряде случаев этот опыт может накапливаться «впрок», «на всякий случай» и найти применение значительно позже в экстренных жизненных ситуациях.
Совершенствование двигательной активности в играх животных
Манипуляционные игры
Признав игру развивающейся деятельностью, необходимо теперь уточнить, что именно и как при этом развивается, что нового привносит игровая активность в поведение животного. Наиболее удобно это сделать при рассмотрении манипуляционных игр, т.е. игр молодых животных с предметами. В качестве примера возьмем игровые манипуляции детенышей хищных млекопитающих (наблюдения Фабри и Мешковой).
Первые действия игрового типа появляются у детенышей животных только после прозрения. У лисенка, например, этот срок наступит на 12-й день после рождения. Уже говорилось о том, что до этого вся его манипуляционная активность сводится всего лишь к нескольким действиям, причем манипуляции, производимые двумя передними конечностями (без участия челюстного аппарата), представлены только одной, наиболее примитивной формой, и еще полностью отсутствуют действия, производимые лишь одной передней лапой.
Когда же после открывания глаз и последующего вскоре за этим выхода из гнезда детеныш начинает играть (у псовых в возрасте 16–23 суток), то это приводит к подлинному скачку в развитии моторной сферы, причем резко увеличивается как число форм манипулирования (у лисенка от 8 до 28), так и число объектов манипулирования.
Появляются «игрушки» – объекты игры. Новые действия детеныша уже не связаны с сосанием, и их отличительная черта состоит в большой общей подвижности молодого животного. Лисята трогают, хватают, обкусывают все, что им попадается, перетаскивают все, что могут, в зубах с места на место, треплют небольшие предметы, зажав их в зубах и мотая головой из стороны в сторону. Типичными и часто выполняемыми формами игрового обращения с предметами являются также поддевание объекта носом (часто с последующим подбрасыванием), удерживание объекта частично или целиком навесу в зубах (в первом случае объект опирается одним концом на субстрат), придерживание объекта ртом или носом на вытянутых вперед передних конечностях, которые неподвижно лежат на субстрате (объект при этом покоится на них, как на подставке), подгребание объекта передними лапами к себе, прижимание объекта к телу, лежа на спине, с одновременным обкусыванием, подталкиванием и передвиганием по поверхности тела носом или передними конечностями. В других случаях объект прижимается конечностями к субстрату, и одновременно часть объекта оттягивается вверх или в сторону зубами. Часто производятся роющие движения и др.
Впервые появляются манипуляции, выполняемые лишь одной передней конечностью. Это прикасание, поглаживание, придавливание объектов одной кистью, прикасание к объектам краем кисти с отводящими или приводящими движениями конечности, притягивание объектов лапой с их защемлением согнутыми пальцами или зацеплением их за края когтями.
Как мы видим, в ювенильном периоде онтогенеза происходит существенное обогащение двигательной активности молодого животного (рис. 22). При этом, однако, новые формы манипулирования строятся преимущественно на основе первоначальных, доигровых форм и представляют собой лишь модификацию первичных форм деятельности на большом числе новых, разнокачественных объектов. Другими словами, качественные изменения в поведении детеныша, сопряженные с началом игровой активности, являются результатом развития доигровых форм манипулирования, созревания моторных и сенсорных компонентов этого первичного манипулирования.
Рис. 22. Развитие манипуляционной активности в онтогенезе у лисицы и барсука. Обозначения: I – от рождения до 2 часов; II – от 2 часов до 2 суток; III – от 2 суток до прозрения (у лисят до 12 суток, у барсучат – до 44); IV – от прозрения до 2 месяцев; V – от 2 до 3 месяцев; VI – взрослое состояние. А – манипуляции, выполняемые ротовым аппаратом и всей головой без участия передних конечностей; Б – манипуляции, выполняемые передними конечностями совместно с ротовым аппаратом; В – манипуляции, выполняемые только обеими передними конечностями; Г – манипуляции, выполняемые только одной передней конечностью. Арабские цифры у столбиков – проценты. (Исследование Фабри и Мешковой)
Так, например, у лисенка возможность взять разные небольшие предметы ртом заранее дана предшествующим хватанием соска, возможность трепания (резкими движениями головы из стороны в сторону) – расталкиванием головой собратьев, приминания и перемещения «игрушек» к субстрату и по нему – соответствующими движениями головой и конечностями в области соска. Даже такие сравнительно сложные формы игрового обращения с предметами, как их обкусывание с одновременным удерживанием передними конечностями на субстрате или удерживание субъекта по рту с одновременным его подталкиванием передними конечностями (лежа на животе или боку), можно без труда вывести из таких первичных пищевых движений, как поиск соска, его захватывание и сосание с одновременным упором в живот матери. Из этих двигательных элементов вырастает и отбрасывание или подбрасывание схваченного предмета в сторону или вверх и т.д.
В целом мы находим во всех игровых действиях (включая и вновь появившиеся действия одной конечностью) проявления расширения и усиления первичных дополнительных функций ротового аппарата и передних конечностей, что стало возможным в результате физического развития детеныша, позволяющего ему вступить в более разнообразные отношения с окружающим миром. Таким образом, по отношению к предыгровым манипуляциям игры молодых животных с предметами представляют собой новые формы манипулирования, которые, однако, состоят из прежних, но функционально усиленных и расширенных моторных элементов. Поэтому игру молодых животных и необходимо признать развивающейся деятельностью.
Следует отметить, что это соотношение между первичной двигательной активностью и последующей имеет место не только в сфере дополнительных функций, о которых до сих пор говорилось, но и в сфере главных функций основных эффекторных систем. Так, вместо одного пищевого объекта (молоко) появляется разнообразная пища, различная по своим физическим качествам. Такую пищу можно употребить лишь после предварительной обработки. И если раньше было достаточно одного пищевого действия – сосания, то сейчас молодому животному приходится овладевать многими новыми формами обращения с пищевыми объектами, тренировать и совершенствовать соответствующие движения, что и происходит во время манипуляционных игр.
Если обратиться к нашему примеру – лисенку, то первоначально главная (пищевая) функция ротового аппарата представлена сосанием. Вскоре после прозревания лисята начинают облизывать пищевые (как и непищевые) объекты (в возрасте до 21 суток), а затем есть твердую пищу. Соответствующие движения челюстного аппарата берут свое начало от его первичных хватательных движений. Поскольку, однако, первичные хватательные движения челюстного аппарата не служили для поедания захватываемого объекта (сосок не откусывается и не съедается), то мы имеем здесь дело со сменой функций.
Как способ приема жидкой пищи появляется лакание, причем характерно, что первоначально лакающие движения весьма напоминают первичные сосательные: язык складывается желобком, через который жидкость втягивается. Постепенно эти движения превращаются в подлинное лакание.
Итак, мы видим, что значение игровой активности в формировании моторных компонентов поведения определяется глубокими качественными преобразованиями в двигательной сфере с проявлениями расширения и интенсификации функции, отчасти субституции (переход некоторых дополнительных функций от ротового аппарата к конечностям или наоборот), а иногда и смены функции. Все это происходит в результате упражнения и совершенствования немногих изначальных моторных элементов, причем количественные изменения последних приводят к появлению качественно новых форм манипулирования.
Биологическая обусловленность манипуляционных игр
До сих пор мы рассматривали манипуляционные игры только на примере одного вида хищных млекопитающих (лисицы), у которого, однако, эта активность развита слабее, чем у многих других представителей этого отряда, особенно если иметь в виду медведей, енотов и кошек. У этих животных отмеченные качественные преобразования еще более разительны, поскольку они обладают намного более мультифункциональными передними конечностями, чем псовые. Большинство манипуляций выполняется лисицей, подобно другим псовым, только челюстным аппаратом, так как псовые обладают олигофункциональными («олиго» – мало) конечностями, специализированными к быстрому длительному бегу. В этих условиях ротовой аппарат сохраняет в большой степени дополнительные двигательные функции, которые, например, у медведей присущи передним конечностям. Поэтому субституционные преобразования, равно как явления расширения, усиления и смены функции, выступают в манипуляционных играх медвежат значительно интенсивнее и разнообразнее, чем у лисят (Фабри).
По-иному проявляется в играх специализация передних конечностей у барсука. По данным Мешковой, у барсучат эти конечности больше участвуют в игровых действиях, чем у детенышей псовых. Это связано со специализацией барсука к норному образу жизни и питанию относительно малоподвижной пищей. В жизни взрослых барсуков большую роль играют такие формы манипулирования, как рытье и транспортировка грунта передними конечностями, сгребание ими подстилочного материала и т.д. Эти действия и развиваются в играх барсучат с предметами.
Весьма однообразными являются манипуляционные игры у детенышей копытных. Немногочисленные манипуляции, выполняемые головой или передними конечностями, состоят в толкании (носом), нанесении ударов, кусании, скребущих и роющих движениях. Совершенно отсутствуют манипуляции, выполняемые совместно челюстным аппаратом и конечностями или одновременно обеими передними конечностями. Это, как и вообще весь характер игры у детенышей копытных, отражает специфическую обусловленность игр образом жизни этих животных. Предельная специализация двигательного аппарата к основной, опорно-локомоторной, функции сходит к минимуму способность манипулировать.
Диаметрально противоположная картина наблюдается у обезьян. У этих животных грудные (передние) конечности наименее специализированы, и их дополнительные функции получили предельное развитие среди млекопитающих (и вообще всех животных). Соответственно этому мы находим в играх молодых обезьян не только намного больше двигательных элементов, чем у других животных, но и качественно новые формы, о которых еще пойдет речь.
Видотипичные особенности поведения, обусловленные образом жизни, достаточно четко проявляются уже в пределах одного семейства. Так, в играх лисят появляются типичные для лисиц внезапные высокие прыжки («мышкование» взрослых особей), как и элементы характерных для взрослых особей приемов борьбы и охоты (специфические формы подкарауливания, подкрадывания, нападения, умерщвления жертвы и т.п.). С другой стороны, по Мешковой, игры щенков динго более однообразны, чем игры лисят. В них преобладает длительное преследование друг друга, в чем воплощается такая характерная особенность поведения взрослого динго, как охота гоном (стремительным преследованием жертвы).
Особенно заметно на игровой активности, в частности на сроках появления манипуляционных игр, сказывается степень зрелорождения детенышей, но сущность процесса от этого не меняется.
Ювенильное манипулирование и взрослое поведение
Двигательный репертуар взрослого животного, как нам уже известно, формируется путем обрастания и дополнения инстинктивной, врожденной основы поведения видотипичным индивидуальным опытом, т.е. путем облигатного научения. Теперь можно добавить, что у высших животных это облигатное научение совершается в ювенильном периоде, перед половым созреванием, в форме игровой активности. При этом имеется в виду, что, как и в раннем постнатальном периоде, облигатное научение включает в себя также факультативные компоненты или сочетается с таковыми.
Этот процесс, как только что было показано, знаменуется модификацией первичных доигровых двигательных элементов. В ходе игры из них формируются, упражняются и совершенствуются существенные компоненты поведения взрослого животного. Это выражается и в том, что с возрастом все усложняющееся манипулирование приобретает все больше видотипичных черт.
Модификация доигровых двигательных элементов происходит по мере того, как предметная деятельность молодого животного распространяется на все новые объекты, в связи с чем растет моторно-сенсорный опыт животного, устанавливаются все более важные связи с биологически значимыми компонентами среды, в том числе с сородичами (о формировании группового поведения см. ниже).
При таком понимании игры как развивающейся деятельности становится ясным, почему игровые действия выполняются «не всерьез». Ведь «серьезные» действия являются завершением этого развития. В полной мере выявляется и биологический эффект игр, в данном случае манипуляционных, который выражается в построении, совершенствовании, «отработке» моторных (и сенсорных) компонентов поведенческих актов. Подчеркнем еще раз, что игры молодых животных – это не действия, аналогичные таковым взрослых животных, а сами эти «взрослые действия» на стадии их формирования из более примитивных морфофункциональных элементов.
Формирование общения в играх животных
Совместные игры
Групповое поведение у высших животных также формируется в большой степени в процессе игры. Эту роль выполняют совместные игры: под ними следует понимать такие игры, при которых имеют место согласованные действия хотя бы двух партнеров. Совместные игры встречаются только у животных, которым свойственны развитые формы группового поведения.
Конечно, общение формируется не только в ходе совместной игровой активности детенышей. Достаточно указать на то, что ритуализованные формы поведения, эти важнейшие компоненты общения, в полной мере проявляются и у таких животных, которые с момента рождения выращивались в полной изоляции и не имели никакой возможности общаться, а тем более играть с другими животными. К генетически фиксированным, инстинктивным формам общения относится и взаимное стимулирование (аллеломиметическое поведение). Что же в таком случае формируется в сфере общения у высших животных лишь в результате игровой активности? Чтобы ответить на этот вопрос, необходимо и в данном аспекте проанализировать игровую активность молодых животных как развивающуюся деятельность, учитывая при этом, что и общение является одной из форм деятельности животных.
Совместные игры выполняются преимущественно без предметов. Как и в манипуляционных играх, в них проявляются особенности образа жизни животного. Так, среди грызунов у детенышей морской свинки отсутствует игровая борьба, и их игры ограничиваются совместными прыжками и боданием, которое служит «приглашением» к игре. Стычки приводят у этих животных всегда к повреждениям, но первые драки появляются лишь одновременно с подлинным половым поведением, т.е. на 30-й день жизни. У сурка же, например, ведущим проявлением игровой активности является именно игровая борьба: молодые животные часто и подолгу «борются», поднимаясь на задние конечности и обхватывая друг друга передними. В такой позе они трясутся и толкаются. Часто наблюдается у них и игровое бегство, в то время как общеподвижные игры встречаются у молодых сурков редко.
Широко распространена игровая борьба у хищных. У куньих преобладают охотничьи игры (помимо общеподвижных), которые нередко переходят в игровую борьбу. Немецкий этолог К.Вюстехубе видит в этом подготовку к борьбе с крупной, мощно сопротивляющейся добычей. Как и у других млекопитающих, преследователь и преследуемый в таких играх часто меняются ролями. У медвежат игровая борьба выражается в том, что партнеры толкают и «кусают» друг друга, обхватившись передними лапами, или наносят друг другу удары. Встречаются также совместные пробежки (или плавание наперегонки), игра в «прятки» и т.п. Интересно отметить что, как показали полевые наблюдения исследователя поведения медведей П.Кротта, детеныши бурого медведя играют в естественных условиях реже, чем детеныши других хищных. Кротт объясняет это тем, что мать с самого начала не приносит медвежатам корм и им приходится тратить много времени на его поиск, поэтому играть зачастую «некогда».
Немецкому этологу Р. Шенкелю удалось на основе двухлетних полевых исследований, проведенных в Кении, проследить формирование игрового поведения львят с первых дней их жизни. Их совместные игры (как и у других детенышей кошачьих) заключаются прежде всего в подкрадывании, нападении, преследовании, и «борьбе», причем партнеры то и дело меняются ролями. Шенкель подчеркивает, что игра львят служит формированию взрослого поведения, но первоначально она состоит только из диффузных двигательных элементов. Это еще одно подтверждение концепции игры как развивающейся деятельности.
Отношения, складывающиеся между партнерами в ходе совместных игр, особенно при игровой борьбе, принимают чаще всего иерархический характер. Элементы антагонистического поведения и соподчинения были установлены в играх многих млекопитающих. Правда не всегда эти субординационные отношения непосредственно перерастают в таковые у взрослых особей.
У псовых иерархические отношения начинают формироваться в возрасте 1–1,5 месяца, хотя соответствующие выразительные позы и движения появляются в ходе игры уже раньше. Так, у лисят уже на 32–34-й день жизни наблюдаются вполне выраженные «нападения» на собратьев с признаками импонирования и запугивания. Уже в начале второго месяца жизни появляются иерархические отношения у койотов. Темброк показал, что в отличие от взрослых особей отношения господства и подчинения возникают у детенышей псовых на основе неритуализованного агрессивного общения. Следовательно, ритуализованные формы общения хотя и появляются в онтогенезе довольно рано, начинают выполнять свою роль значительно позже.
Что же касается неритуализованного игрового общения молодых животных, в частности псовых, то, вероятно, все их движения также имеют сигнальное значение. Так, например, «трепание» партнера – не только проявление «грубой» физической силы, оно носит признаки и неритуализованного демонстрационного поведения, являясь средством психического воздействия на партнера, запугивания. Аналогично обстоит дело и с наскакиванием на партнера и т.п.
В отличие от рассмотренных до сих пор совместных неманипуляционных игр в совместных манипуляционных играх животные включают в свои совместные действия какие-нибудь предметы в качестве объекта игры, поэтому общение между партнерами носит при таких играх отчасти опосредованный характер. Такие игры выполняют большую коммуникативную роль, хотя подобные предметы могут одновременно служить и заменой натурального пищевого объекта (у хищных – жертвы).
В качестве примера совместной манипуляционной игры можно привести описанные Вюстехубе совместные действия трех молодых хорьков с пустой консервной банкой. Будучи случайно уроненной в таз умывальника, эта банка затем многократно сбрасывалась ими туда, что производило соответствующий шумовой эффект. Когда же зверькам вместо банки давался резиновый мячик, хорьки с ним так не играли, но впоследствии находили другой твердый предмет – фаянсовую затычку, с помощью которой возобновляли ту же «шумовую» игру.
У диких четырехмесячных поросят немецкий этолог Г.Фрэдрих однажды наблюдал оживленную совместную игру с монетой: поросята нюхали и придавливали ее «пятачками», подталкивали, хватали зубами и подбрасывали ее вверх, резко вскидывая при этом голову. В этой игре одновременно участвовало несколько поросят, причем каждый из них старался овладеть монетой и сам поиграть с ней описанным образом. Фрэдрих также наблюдал совместные игры молодых кабанов с тряпками. Подобно щенкам, поросята хватали зубами одновременно одну и ту же тряпку и тянули ее в разные стороны. «Победитель» или убегал с тряпкой, или продолжал самостоятельно с ней играть, трепал ее и т.д.
В подобных «трофейных» играх отчетливо выступают и элементы демонстрационного поведения, причем достигается импонирующий эффект с помощью объекта – «посредника», точнее, путем демонстрации его обладания. Не меньшую роль играют, конечно, «оспаривание», захват, отнятие объекта, равно как непосредственная «проба сил», когда животные, схватившись одновременно за объект, тянут его в разные стороны.
Игровая сигнализация
Согласованность деятельности игровых партнеров основывается на обоюдной врожденной сигнализации. Эти сигналы выполняют функцию ключевых стимулов игрового поведения. Это – специфические позы, движения, звуки, оповещающие партнера о готовности к игре и «приглашающие» его принять в ней участие. Так, например, у бурого медведя, по полевым наблюдениям Кротта, «приглашение» к игре состоит в том, что медвежонок медленно приближается к возможному игровому партнеру, качая головой влево и вправо, потом припадает к земле и очень осторожно обхватывает его передними лапами. У детенышей псовых «приглашение» к игре осуществляется с помощью особой («игровой») манеры приближения к партнеру, специфическим раскачиванием головы из стороны в сторону, пригибанием книзу передней части туловища, сопровождающимся его раскачиванием или небольшими прыжками из стороны в сторону на виду у партнера, поднятием передней лапы в сторону партнера и т.п. У «заигрывающего» детеныша одновременно появляются продольные складки на лбу, а ушные раковины обращены вперед. У виверр «заигрывание» происходит посредством наскакивания, легкого покусывания и особенно прыжков вверх, у обезьян – путем наскакивания, подергивания за хвост или волосы, шлепков и т.д.
Не менее важными являются сигналы, предотвращающие «серьезный» исход игровой борьбы, позволяющие животным отличить игру от «не игры». Без подобного предупреждения о том, что агрессия «ненастоящая», игровая борьба может легко перейти в подлинную. Эти сигналы явно родственны позам и движениям «умиротворения» при подлинных стычках взрослых животных, они главным образом и создают общую «игровую ситуацию».
Принципиальный интерес представляет то обстоятельство, что сигналы «заигрывания», «приглашения к игре», по существу, являются сигналами подлинного агрессивного нападения, и только в игровой ситуации они функционируют как «мирные». Это показывает, что и в сфере общения игровое поведение характеризуется явлениями смены функции подобно тому, как мы это отмечали в отношении развития двигательных способностей. Аналогичным образом в игровом общении обнаруживаются и явления рекомбинации врожденных сигналов или их элементов, совершенствование их сочетания и применения, т.е. имеют место расширение и интенсификация функции, а также явления субституции функции. В этом нет ничего неожиданного, ибо общение развивается при игровой активности в единстве с моторикой.
Важно еще отметить, что коммуникация осуществляется у животных не только с помощью специальных сигнальных движений. Наоборот, едва ли можно в поведении молодого животного выделить какие-то движения, которые были бы абсолютно лишены сигнального значения. Очевидно, можно говорить лишь о различном удельном весе сигнального компонента двигательных актов. Максимальный вес этот компонент имеет, конечно, в ритуализованных движениях, которые уже всецело служат для коммуникации.
Значение совместных игр для взрослого поведения
Для многих видов животных доказано, что если детенышей лишить возможности совместно играть, то во взрослом состоянии сфера общения окажется заметно ущемленной или даже искаженной. Так, у морских свинок это выражается в сохранении инфантильного поведения, даже после полного полового созревания, и в ненормальных реакциях на сородичей и других животных (П. и И. Кункель). Самцы крыс нуждаются для выполнения воспроизводительной функции в раннем игровом общении с другими крысятами. В этих играх содержатся основные двигательные элементы взрослого самцового поведения. У норок самцы научаются нормальному общению с брачным партнером в ходе совместных игр начиная с 10-недельного возраста; щенки койота, выращенные без игрового общения, обнаруживали повышенную агрессивность и т.д.
Особенно отчетливо значение совместных игр детенышей для дальнейшей жизни особи проявляется у обезьян. О пагубных последствиях лишения молодых обезьян возможности играть со сверстниками (или другими животными) убедительно свидетельствуют эксперименты многих исследователей, в частности Харлоу и его сотрудников. Как и у других животных, обусловленные этим нарушения обнаруживаются у взрослых особей прежде всего в их неспособности к нормальному общению с себе подобными, особенно с половыми партнерами, а также в материнском поведении. Характеризуя роль игры в развитии общения у обезьян, известные исследователи поведения, приматов С. Л. Уошберн и И. де Воре подчеркивали, что без игры невозможно развитие нормальных форм общения и стадного поведения в целом. Молодые обезьяны учатся общаться друг с другом в «игровых группах», т.е. группах совместно играющих детенышей, где они, по Уощберн и де Воре, «практикуют умения и формы поведения взрослой жизни».
Важно отметить, что функцию партнера по игре с успехом может осуществлять другое животное или даже, человек. Об этом свидетельствует, например, то обстоятельство, что при изолированном выращивании детенышей обезьян, имевших возможность играть только с людьми, формирование полноценных форм общения происходит без помех. Аналогично обстоит дело и у детенышей хищных млекопитающих, в частности медвежат и волчат, когда воспитывающий их человек или другие животные (например, собака) заменяют естественных игровых партнеров. Все эти животные в дальнейшем оказываются вполне способными к нормальному общению с сородичами.
Познавательная функция игровой активности животных
Игра и исследовательское поведение
В ходе игры молодое животное приобретает разнообразную информацию о свойствах и качествах предметов в окружающей его среде. Это позволяет конкретизировать, уточнять и дополнять накопленный в процессе эволюции видовой опыт применительно к конкретным условиям жизни особи.
В трудах ряда авторитетных ученых отмечается связь игры с исследовательской деятельностью (Гроос, Бич, Ниссен, Лоренц и др.), но при этом отмечаются и различия между этими категориями поведения. Возражая против взгляда на игру как на «игру природы», якобы не имеющую значения для сохранения вида, Лоренц подчеркивал ее большое значение для «исследовательского научения», ибо в ходе игры животное относится практически к каждому незнакомому предмету как к потенциально биологически значимому и тем самым выискивает в самых различных условиях возможности для существования. Особенно это относится, по Лоренцу, к таким «любопытствующим существам» («Neugierwesen»), как врановые птицы или крысы, которые благодаря чрезвычайно развитому исследовательскому поведению сумели стать космополитами. Аналогичным образом видный немецкий этолог О.Келер указывал на то, что игра является «практически непрестанным поиском проб и ошибок», в результате чего животное медленно, случайно, но подчас и внезапно научается тому, что весьма важно для него.
Правда, другие специалисты высказывают мнение, что сходство между явлениями игры и исследовательского поведения является лишь внешним и не имеет существенного значения. Такой точки зрения придерживаются, например, Хамильтон и Марлер. Однако никем не ставилось под сомнение, что приобретение информации путем игры осуществляется по меньшей мере в сочетании с «собственно» исследовательской деятельностью. Безусловно, не всякая ориентировочно-исследовательская деятельность является игрой, как и ознакомление с окружающим осуществляется у молодого животного не только в игровой форме. Но в каждой игре содержится в той или иной степени исследовательский компонент.
Особенно это относится к играм с предметами, к манипуляционным играм, но опять-таки не всякое манипулирование является игрой. (Не является, например, игрой манипулирование пищевыми объектами во время еды или гнездостроительным материалом во время сооружения гнезда.) Но манипулирование «биологически нейтральными» объектами или же биологически значимыми, но вне их адекватного применения является ни чем иным, как игрой.
Важно подчеркнуть далее, что всякое манипулирование, особенно игровое, всегда включает в себя исследовательский компонент. Более того, манипулирование «биологически нейтральными» предметами представляет собой высшую форму ориентировочно-исследовательской деятельности. С другой стороны, без игры молодое животное может ознакомиться со свойствами только объектов, имеющих для него непосредственно биологическое значение. Игровое манипулирование предметами особенно стимулируется появлением новых или малоизвестных объектов. Роль новизны предметных компонентов среды в манипулировании особенно подчеркивал для обезьян еще Войтонис.
Развитие двигательных способностей всегда сопряжено с исследованием окружающей среды. Можно сказать, что всевозрастающее приобретение информации о компонентах среды является функцией развивающейся двигательной активности, ориентация которой во времени и пространстве в свою очередь осуществляется на основе этой информации. Именно в этом находит свое выражение единство моторных и сенсорных элементов поведения, развивающихся в ходе игры.
В наименьшей мере исследовательский компонент представлен в играх, служащих лишь своего рода «физическими упражнениями»; в наибольшей же мере – там, где имеет место активное воздействие на объект игры, особенно деструктивного порядка, т.е. в манипуляционных играх. Последние могут в некоторых случаях приобрести значение подлинных «исследовательских» игр (см. ниже).
Особое место занимают опосредованные игры, в частности «трофейные» игры, когда, очевидно, можно говорить даже о совместном познавании объекта игры при совместных двигательных упражнениях. Однако эти игры все же служат прежде всего средством общения между животными и установлению определенных отношений между ними, как это имеет место и при других совместных играх. К тому же нельзя, конечно, быть уверенным, что при «трофейных» играх партнеры действительно воспринимают структурные изменения в объекте игры как таковые, ибо их внимание устремлено друг на друга.
Инстинктивные основы игрового познавания
В начале постнатального онтогенеза врожденное узнавание и запечатление служат для первичной ориентации и срочного накопления наиболее необходимого для особи индивидуального опыта. Однако, как уже указывалось, и на протяжении дальнейшей жизни особи врожденное узнавание и запечатление не теряют своего значения, хотя, разумеется, и проявляются по-иному. Запечатление, например, играет важную роль в формировании родительского поведения («обратное запечатление»), врожденное узнавание же выступает на протяжении жизни особи в разных формах как составная часть инстинктивного поведения (узнавание ключевых раздражителей).
В ювенильном периоде, как тоже уже отмечалось, упомянутые врожденные компоненты в основном сливаются с игровой активностью, образуя ее инстинктивную основу. При этом накопление сугубо индивидуального (факультативного) опыта переплетается с видотипичным, инстинктивным приобретением информации, основанным на врожденном узнавании.
Врожденное узнавание дает играющему животному прежде всего возможность узнать о пригодности определенного предмета или индивида к включению его в игру, руководствуясь при этом соответствующими ключевыми раздражителями. Последние представляют собой пусковые стимулы партнера или определенные свойства замещающего объекта, воспринимаемые в самом обобщенном виде (например, само движение объекта игры). Так, Лоренц приводит следующие признаки, которыми должен обладать объект охотничьей игры котенка в качестве ключевых раздражителей: маленькое, округлое, мягкое, все, что быстро движется, катаясь или скользя, и, главное, все, что «убегает». И в ходе самой игры с партнером или заменяющим его предметом молодое животное реагирует на ключевые раздражители, управляющие упражнением и развитием соответствующих инстинктивных действий.
Можно, очевидно, сказать, что если иметь в виду упражнение и развитие инстинктивных действий, то молодому животному, по сути дела, «все равно», с чем и кем играть, лишь бы объект игры или игровой партнер обладал соответствующими ключевыми раздражителями.
Если же, наоборот, иметь в виду познавательную ценность игры, то именно эти ключевые раздражители приводят молодое животное к биологически наиболее важным для него компонентам среды и обеспечивают тем самым всестороннее ознакомление с ними. Иными словами, врожденное узнавание является здесь необходимой предпосылкой познания носителей ключевых раздражителей уже на перцептивном уровне. Говоря словами Лоренца, путем игры котенок научается узнавать, «что такое мышь». Речь идет здесь именно о мыши как таковой, а не о совокупности присущих ей пусковых стимулов (маленькое, округлое и т.д.), которые воспринимаются животным как ощущения путем врожденного узнавания.
Расширение функции в игровом познавании
При переходе компонентов доигрового поведения в ювенильный период обогащение и трансформация первичных элементов исследовательского поведения совершаются в процессе игры по тем же закономерностям, что и развитие двигательных и коммуникативных компонентов раннего постнатального поведения.
В качестве примера можно указать на расширение функции в сфере исследовательского поведения у незрелорождающихся млекопитающих, в частности хищных. На начальном этапе постнатального развития детеныша получаемая им в гнезде информация является минимальной. С началом же игрового этапа двигательная активность детеныша претерпевает глубокие преобразования, охарактеризованные выше; начинают функционировать дистантные рецепторы, начинается полноценное общение с матерью и собратьями. Все это коренным образом изменяет и, главное, обогащает воспринимаемую детенышем информацию об окружающей его среде. Наконец, с выходом детеныша из гнезда вновь наступает коренное, на этот раз решающее, изменение его двигательной, коммуникативной и соответственно познавательной деятельности. Все его поведенческие акты выполняются уже в совершенно новых условиях: объектами воздействия являются уже не только материнская особь, собратья и немногочисленные предметы внутри гнезда, но ими становятся прежде всего многие разнокачественные предметы с различной биологической валентностью. А это установление новых связей с компонентами среды обеспечивает поступление мощного потока разнообразной жизненно необходимой информации. Огромную роль играют и появляющиеся в это время индивидуальные игры с предметами, доставляющие молодому животному особенно много разнообразных сведений о потенциально биологически значимых объектах. Именно к этой категории игры относятся в первую очередь и манипуляционные игры с «биологически нейтральными» предметами.
Итак, в ходе онтогенеза все больше расширяется и усложняется познавательная деятельность развивающегося животного, имеет место типичное расширение функции, а поскольку исследовательское поведение после выхода из гнезда обращается на качественно совершенно новые объекты, отчасти можно говорить и о явлениях смены функции.
Высшие формы игровой исследовательской деятельности животных
При всем многообразии форм игры их объединяет большая общая подвижность животного, большое разнообразие производимых им телодвижений и интенсивное перемещение в пространстве (рис. 23). Наиболее ярко это выступает, конечно, в играх, которые носят чисто локомоторный характер, выражаясь в разных формах интенсивного передвижения, или которые направлены на собственное тело (игра с собственным хвостом и т.п.). Однако, как мы видели, и в других категориях игровой активности развиваются прежде всего двигательно-сенсорные координации (например, глазомер) и общие физические способности (ловкость, быстрота, реактивность, сила и т.п.). Одновременно упражняются определенные элементы поведения в функциональных сферах питания, защиты и нападения, размножения и др., совершенствуются и развиваются средства общения, устанавливаются отношения с сородичами, причем иногда в виде иерархических взаимоотношений. При всем этом происходит рекомбинация элементов доигрового поведения, в результате чего формируются и совершенствуются новые проявления видотипичного, инстинктивного поведения на более высоком уровне. Как было показано, игровое поведение направляется ключевыми раздражителями независимо от их носителя, но одновременно животное приобретает жизненно важную информацию об этих носителях, об их внешнем виде и о некоторых их физических качествах (вес, прочность, подвижность и т.п.). Однако в целом при всех рассмотренных до сих пор играх происходит лишь поверхностное ознакомление с компонентами среды, чем и ограничивается познавательный компонент этих игр.
У молодых обезьян (у низших обезьян преимущественно в возрасте 2–5 лет) наблюдаются манипуляционные игры совсем иного рода, которые необходимо признать играми высшего типа (Фабри). В противоположность рассмотренным играм этот тип игры характеризуется прежде всего сложными формами обращения с предметами при незначительной общей подвижности животного: лишь изредка меняя свое местонахождение, животное подолгу и сосредоточенно манипулирует предметом, подвергает его разнообразным, преимущественно деструктивным воздействиям или даже воздействует им на другие объекты. В последнем случае иногда выполняются манипуляции, сходные с орудийными действиями взрослых обезьян (рис. 24).

Рис. 23. Игра детеныша обезьяны (макака-резуса): раскачивание на ветке, случайно застрявшей в петлях сетчатой стенки, когда детеныш бегал, прыгал и лазал с ней по клетке (опыты Фабри)
Рис. 24. Манипуляционные игры молодых низших обезьян с проволочками (пример игры высшего типа). А–И – игры с железной проволокой: А – царапанье субстрата концом проволоки, Б– всовывание проволоки в отверстие и надавливание на свободный конец пальцами, В– «вплетение» проволоки в сетчатую стенку клетки, Г–И– вкладывание проволоки (Ж) в щель полки и последующее манипулирование ею под полкой; К–О – игра с алюминиевой проволокой, случайно принявшей вид крючка в результате интенсивного обкусывания: НК–вид «крючка», Л– О–просовывание проволоки сквозь сетку с последующим протягиванием ее (исследование Фабри)
При подобных сложных играх с предметами совершенствуются высокодифференцированные и тонкие эффекторные способности (прежде всего пальцев) и развивается комплекс кожно-мышечной чувствительности и зрения. Познавательный аспект приобретает здесь особую значимость: животное обстоятельно и углубленно знакомится со свойствами предметных компонентов среды, причем особое значение приобретает исследование внутреннего строения объектов манипулирования в ходе их деструкции. Особое значение приобретает и то обстоятельство, что объектами манипулирования являются чаще всего «биологически нейтральные» предметы. Благодаря этому существенно расширяется сфера получаемой информации: животное знакомится с весьма различными по своим свойствам компонентами среды и приобретает при этом большой запас разнообразных потенциально полезных «знаний». Наличие таких игр у обезьян, несомненно, связано с их отличительными по сравнению с другими животными психическими способностями, в частности их «ручным мышлением», о чем еще пойдет речь.
Таким образом, здесь имеют место подлинные исследовательские, познавательные игры, причем игры высшего порядка. Выполняются эти игры в одиночку. Помимо общего накопления сведений «про запас» такие познавательные игры также прямо и непосредственно готовят животное к взрослому поведению (например, в сфере питания, где дифференцированные движения пальцев играют немалую роль при расчленении плодов, выковыривании семян и т.д.). Включаются такие игры и в сферу общения. Однако так как познавательная функция играет здесь ведущую роль, эти игры приобретают характер самостоятельной сферы поведения с собственным функциональным (специально-познавательным) значением.
Вместе с тем и в этих наиболее сложных проявлениях игровой активности, которые пока удалось обнаружить только у высших приматов, обнаруживается инстинктивная основа, этологические критерии которой вполне соответствуют критериям типичной функциональной сферы (автохтонность, замещение активности, супероптимальное реагирование и т.д.). Это вновь показывает, что общая закономерность единства врожденного и благоприобретаемого поведения сохраняет свое значение в полной мере на всем протяжении онтогенеза, в том числе и на высших филогенетических уровнях.
Часть III
ЭВОЛЮЦИЯ ПСИХИКИ
Вводные замечания
Эволюция психики составляет часть общего процесса эволюции животного мира и совершалась по закономерностям этого процесса. Повышение общего уровня жизнедеятельности организмов, усложнение их взаимоотношений с окружающим миром приводило в ходе эволюции к необходимости все более интенсивного контактирования со все большим числом предметных компонентов среды, к совершенствованию передвижения среди этих компонентов и ко все более активному обращению с ними. Только такое прогрессивное изменение поведения могло обеспечить возрастающее потребление необходимых для жизни компонентов среды, равно как все более успешное избегание неблагоприятных или вредных воздействий и опасностей. Однако для всего этого требовалось существенное совершенствование ориентации во времени и пространстве, что и достигалось прогрессом психического отражения.
Таким образом, движение (первично локомоция, а впоследствии и манипулирование) являлось решающим фактором эволюции психики. С другой стороны, без прогрессивного развития психики не могла бы совершенствоваться двигательная активность животных, не могли бы производиться биологически адекватные двигательные реакции и, следовательно, не могло бы быть эволюционного развития.
Конечно, психическое отражение не оставалось неизменным в ходе эволюции, а само претерпевало глубокие качественные преобразования, к рассмотрению которых мы сейчас и перейдем. Существенным является то, что первично; еще примитивное, психическое отражение обеспечивало только уход от неблагоприятных условий (как это характерно и для современных простейших); поиск непосредственно не воспринимаемых благоприятных условий появился значительно позже. Как мы уже видели, такой поиск является постоянным компонентом развитого инстинктивного поведения.
Равным образом только на высоких уровнях эволюционного развития, когда уже существует предметное восприятие и сенсорные действия животных обеспечивают возникновение образов, психическое отражение становится способным полноценно ориентировать и регулировать поведение животного с учетом предметности компонентов внешней среды. Такое отражение имеет, в частности, первостепенное значение для выделения преград (в узком и широком смысле слова), что, как мы видели, является необходимым условием для появления самых лабильных форм индивидуального поведенческого приспособления к меняющимся условиям среды – навыков и, у наиболее высокоразвитых животных, интеллекта.
Признаки наиболее глубоких качественных изменений, которые претерпела психика в процессе эволюции животного мира, Леонтьев положил в основу выделенных им стадий психического развития. Четкая, наиболее существенная грань проходит между элементарной сенсорной и перцептивной психикой, знаменуя собой основную веху грандиозного процесса эволюции психики. Поэтому в дальнейшем изложении мы будем основываться на этом делении.
Элементарную сенсорную психику Леонтьев определяет как стадию, на которой деятельность животных «отвечает тому или иному отдельному воздействующему свойству (или совокупности отдельных свойств) в силу существенной связи данного свойства с теми воздействиями, от которых зависит осуществление, основных биологических функций животных. Соответственно отражение действительности, связанное с таким строением деятельности, имеет форму чувствительности к отдельным воздействующим свойствам (или совокупности свойств), форму элементарного ощущения».* Стадия же перцептивной психики, по Леонтьеву, «характеризуется способностью отражения внешней объективной действительности уже не в форме отдельных элементарных ощущений, вызываемых отдельными свойствами или их совокупностью, но в форме отражения вещей».** Деятельность животного определяется на этой стадии тем, что выделяется содержание деятельности, направленное не на предмет воздействия, а на те условия, в которых этот предмет объективно дан в среде. «Это содержание уже не связывается с тем, что побуждает деятельность в целом, но отвечает специальным воздействиям, которые его вызывают».***
* Леонтьев А. Н. Проблемы развития психики. М., 1959. С. 162–163.
** Там же. С. 176.
*** Там же.
Однако для наших целей этого подразделения недостаточно, и поэтому следует как в пределах элементарной сенсорной, так и в пределах перцептивной психики выделить существенно различающиеся уровни психического развития: низший и высший, допуская при этом существование и промежуточных уровней (рис. 25). Важно отметить, что крупные систематические таксоны животных не всегда и не вполне укладываются в эти рамки. Это неизбежно, так как в пределах крупных таксонов (в данном случае подтипов или типов) всегда имеются животные, стоящие на смежных уровнях психического развития. Объясняется это тем, что качества высшего психического уровня всегда зарождаются на предшествующем уровне.
Кроме того, расхождения между психологической и зоологической классификациями обусловлены тем, что морфологические признаки, на которых построена систематика животных, отнюдь не всегда определяют особенности и степень развития психической деятельности последних. Поведение животных представляет собой совокупность функций эффекторных органов животных. А в процессе эволюции именно функция первично определяет форму, строение организма, его систем и органов.
Рис. 25. Родословное дерево и уровни психического развития животных (по Фабри): 1 – простейшие: II – губки; III – кишечнополостные; IV– плоские черви; V– круглые черви;
VI –кольчатые черви; VII –членистоногие; VIII –моллюски; IX – иглокожие; Х – позвоночные. I – первичные жгутиковые; 2 – современные жгутиковые; 3 – корненожки (амебы); 4 – инфузории; 5– гидроидные; 6 – медузы; 7– ленточные черви; 8 – ресничные черви (планарии); 9 – малощетинковые черви; 10 – пиявки; II – многощетинковые черви; 12 – ракообразные: 13 – паукообразные; 14 – многоножки; 15 –насекомые; 16– двустворчатые моллюски; 17 – брюхоногие моллюски; 18 – головоногие моллюски; 19 – морские лилии; 20 – морские звезды; 21 – оболочники; 22 – бесчерепные (ланцетник); 23 – круглоротые; 24 – рыбы; 25 – земноводные; 26 – пресмыкающиеся; 27 – птицы; 28 – млекопитающие
И лишь вторично строение эффекторов, их двигательные возможности определяют характер поведения животного, ограничивают сферу его внешней активности.
Этот диалектический процесс, однако, осложняется еще возможностями многопланового решения задач (по меньшей мере у высших животных) и компенсаторными процессами в области поведения. Это означает, что если животное в данных условиях лишено возможности решить биологически важную задачу одним путем, оно, как правило, имеет в своем распоряжении еще другие, резервные возможности. Так, одни эффекторы могут замениться другими, т.е. разные морфологические структуры могут служить для выполнения биологически однозначных действий. С другой стороны, одни и те же органы могут выполнять разные функции, т.е. осуществляется принцип мультифункциональности. Особенно гибки морфофункциональные отношения в координационных системах, прежде всего в центральной нервной системе высших животных.
Итак, с одной стороны, образ жизни определяет развитие приспособлений в эффекторной сфере, а с другой стороны, функционирование эффекторных систем, т.е. поведение, обеспечивает удовлетворение жизненных потребностей, обмена веществ в ходе взаимодействия организма с внешней средой. Изменения условий жизни порождают необходимость изменения прежних эффекторных функций (или даже появления новых функций), т.е. изменения поведения, а это затем приводит к соответствующим морфологическим изменениям в эффекторной и сенсорной сферах и в центральной нервной системе. Но не сразу и даже не всегда функциональные изменения влекут за собой морфологические. Более того, у высших животных зачастую вполне достаточными, а иногда даже наиболее результативными являются чисто функциональные изменения без морфологических перестроек, т.е. адаптивные изменения только поведения (А.Н.Северцов). Поэтому поведение в сочетании с мультифункциональностыо эффекторных органов обеспечивает животным наиболее гибкую адаптацию те новым условиям жизни.
Указанные функциональные и морфологические преобразования определяют качество и содержание психического отражения в процессе эволюции. О них, о диалектических взаимоотношениях между образом жизни (биологией) и поведением, необходимо всегда помнить, когда мы говорим о примате двигательной активности, деятельности животных, в развитии психического отражения.
Уместно еще раз напомнить и о том, что, вопреки еще бытующему представлению, врожденное и приобретаемое поведение не являются последовательными ступенями на эволюционной лестнице, а развиваются и усложняются совместно, как два компонента одного единого процесса. Следовательно, не было и нет такого положения, чтобы у низших животных имелись лишь одни инстинкты (или даже рефлексы), на смену которым у высших приходят навыки, а инстинкты все больше сходят на нет. На самом деле, как уже было показано, прогрессивному развитию именно инстинктивного, генетически фиксированного поведения соответствует прогресс в области индивидуально-изменчивого поведения. Инстинктивное поведение достигает наибольшей сложности как раз у высших животных, и этот прогресс влечет за собой развитие и усложнение у этих животных форм научения.
Мы здесь можем, разумеется, дать только самый общий обзор эволюции психики, и то лишь по некоторым его направлениям и на немногих примерах, совершенно не вскрывая всего многообразия путей развития психики в мире животных.
Глава 1 ЭЛЕМЕНТАРНАЯ СЕНСОРНАЯ ПСИХИКА
Низший уровень психического развития
На низшем уровне психического развития находится довольно большая группа животных; среди них встречаются и такие животные, которые стоят еще на грани животного и растительного мира (жгутиковые), а с другой стороны, и сравнительно сложно устроенные одноклеточные и многоклеточные животные. К наиболее типичным представителям рассматриваемой здесь группы животных относятся простейшие, на примере которых здесь будет дана характеристика низшего уровня элементарной сенсорной психики. Правда, некоторые высокоорганизованные простейшие (из числа инфузорий) поднялись уже на более высокий уровень элементарной сенсорной психики. В этой связи следует напомнить, что филогенез простейших шел в известной мере параллельно развитию низших многоклеточных животных, а это, в частности, нашло свое отражение в формировании у простейших аналогов органов таких животных. Эти аналоги получили название органелл.
Движения простейших
Движения простейших отличаются большим разнообразием, причем в этом типе простейших животных встречаются способы локомоции, которые совершенно отсутствуют у многоклеточных животных. Это своеобразный способ передвижения амеб при помощи «переливания» плазмы из одного участка тела в другой. Другие представители простейших, грегарины, передвигаются своеобразным «реактивным» способом – путем выделения из заднего конца тела слизи, «толкающей» животное вперед. Существуют и простейшие, пассивно парящие в воде.
Однако большинство простейших передвигаются активно с помощью особых структур, производящих ритмичные движения, – жгутиков или ресничек. Эти эффекторы представляют собой плазматические выросты, совершающие колебательные, вращательные или волнообразные движения. Жгутиками, длинными волосовидными выростами обладают уже упомянутые примитивные простейшие, получившие свое название благодаря этому образованию. С помощью жгутиков тело животного (например, эвглены) приводится в спиралевидное поступательное движение. Некоторые морские жгутиковые, по данным норвежского ученого И.Трондсена, вращаются при движении вокруг оси со скоростью до 10 оборотов в секунду, а скорость поступательного движения может достигнуть 370 микрон в секунду. Другие морские жгутиковые (из числа динофлягеллят) развивают скорость от 14 до 120 микрон в секунду и больше. Более сложным эффекторным аппаратом являются реснички, покрывающие в большом числе тело инфузории. Как правило, реснитчатый покров располагается неравномерно, реснички достигают на разных участках тела различной длины, образуют кольцевидные уплотнения (мембранеллы) и т.п.
Примером такой сложной дифференциации могут служить инфузории из рода стилонихия. Своеобразные органеллы этих животных позволяют им не только плавать, но и «бегать» по твердому субстрату, причем как вперед, так и назад. Установлено, что координация этих способов и направлений локомоции, как и их «переключение», осуществляется специальными механизмами, локализованными в трех центрах и двух осях градиентов возбуждения в цитоплазме.
Жгутики и реснички приводятся в движение сокращениями миофибрилл, которые образуют волоконца, мионемы, соответствующие мышцам многоклеточных животных. У большинства простейших они являются основным двигательным аппаратом, причем имеются они даже у наиболее примитивных представителей типа жгутиковых. Мионемы располагаются в строгом порядке, чаще всего в виде колец, продольных нитей или лент, а у высших представителей и в виде специализированных систем. Так, у инфузории Caloscolex имеются специальные системы мионем околоротовых мембранелл, глотки, задней кишки, ряд ретракторов отдельных участков тела и т.д.
Интересно отметить, что, как правило, мионемы имеют гомогенную структуру, что отвечает гладкой мускулатуре многоклеточных животных, однако иногда встречаются и поперечно исчерченные мионемы, сопоставимые с поперечно-полосатой мускулатурой высших животных. Все сократительные волоконца служат для выполнения быстрых движений отдельных эффекторов (у простейших – игловидных выростов, щупальцевидных образований и т.п.). Сложные системы мионем позволяют простейшим производить не только простые сократительные движения тела, но и достаточно разнообразные специализированные локомоторные и нелокомоторные движения.
У тех простейших, у которых нет мионем (у амеб, корненожек, споровиков, за одним исключением, и некоторых других простейших), сократительные движения совершаются непосредственно в цитоплазме. Так, при передвижении амебы в наружном слое цитоплазмы, в эктоплазме, происходят подлинные сократительные процессы. Удалось даже установить, что эти явления имеют место всякий раз в «задней» (по отношению к направлению передвижения) части тела амебы.
Таким образом, еще даже до появления специальных эффекторов перемещение животного в пространстве совершается путем сокращений. Именно сократительная функция, носителем которой являются у простейших мионемы, а у многоклеточных мышцы, обеспечивала все разнообразие и всю сложность двигательной активности животных на всех этапах филогенеза.
Кинезы
Локомоция простейших осуществляется в виде кинезов – элементарных инстинктивных движений. Типичным примером кинеза является ортокинез – поступательные движение с переменной скоростью. Если, например, на определенном участке существует температурный градиент (перепад температур), то движения туфельки будут тем более быстрыми, чем дальше животное будет находиться от места с оптимальной температурой. Следовательно, здесь интенсивность поведенческого (локомоторного) акта непосредственно определяется пространственной структурой внешнего раздражителя.
В отличие от ортокинеза при клинокинезе имеет место изменение направления передвижения. Это изменение не является целеустремленным, а носит характер проб и ошибок, в результате которых животное в конце концов попадает в зону с наиболее благоприятными параметрами раздражителей. Частота и интенсивность этих изменений (следовательно, и угол поворота) зависят от интенсивности воздействующего на животное (отрицательного) раздражителя (или раздражителей). С ослаблением силы действия этого раздражителя уменьшается и интенсивность клинокинеза. Таким образом, животное и здесь реагирует на градиент раздражителя, но не увеличением или уменьшением скорости передвижения, как при ортокинезе, а поворотами оси тела, т.е. изменением вектора двигательной активности.
Как мы видим, осуществление наиболее примитивных инстинктивных движений – кинезов – определяется непосредственным воздействием градиентов интенсивности биологически значимых внешних факторов. Роль внутренних процессов, происходящих в цитоплазме, заключается в том, что они дают поведенческому акту «первый толчок», как и у многоклеточных животных.
Ориентация
Уже на примерах кинезов мы видели, что градиенты внешних раздражителей выступают у простейших одновременно как пусковые и направляющие стимулы. Особенно наглядно это проявляется при клинокинезах. Однако изменения положения животного в пространстве еще не являются здесь подлинно ориентирующими, поскольку они носят ненаправленный характер. Для достижения полного биологического эффекта клинокинетические, как и ортокинетические, движения нуждаются в дополнительной коррекции, позволяющей животному более адекватно ориентироваться в окружающей его среде по источникам раздражения, а не только менять характер движения при неблагоприятных условиях.
Ориентирующими элементами являются у представителей рассматриваемого типа и у других низших беспозвоночных, стоящих на данном уровне психического развития, простейшие таксисы. В ортокинезах ориентирующий компонент – ортотаксис – проявляется в изменении скорости передвижения без изменения его направления в градиенте внешнего раздражителя. В клинокинезах этот компонент называется клинотаксисом и проявляется в изменении направления движения на определенный угол.
|
|
|
|
Рис. 26. Клинотаксисное поведение инфузории в температурном градиенте. Попав в теплую зону (вверху), инфузория уходит в более прохладную (внизу) (по Дженнингсу)
Как мы уже знаем, под таксисами понимают генетически фиксированные механизмы пространственной ориентации двигательной активности животных в сторону благоприятных (положительные таксисы) или в сторону от неблагоприятных (отрицательные таксисы) условий среды. Так, например, отрицательные термотаксисы выражаются у простейших, как правило, в том, что они уплывают из зон с относительно высокой температурой воды, реже – из зон с низкой температурой. В результате животное оказывается в определенной зоне термического оптимума (зоне предпочитаемой температуры). В случае ортокинеза в температурном градиенте отрицательный ортотермотаксис обеспечивает прямолинейное удаление от неблагоприятных термических условий. Если же имеет место клинокинетическая реакция (рис. 26), то клинотаксис обеспечивает четкое изменение направления передвижения, ориентируя тем самым случайные клинокинетические движения в градиенте раздражителя (в нашем примере – в термическом градиенте).
Зачастую клинотаксисы проявляются в ритмичных маятникообразных движениях (на месте или при передвижении) или в спиралевидной траектории плывущего животного. И здесь имеет место регулярный поворот оси тела животного (у многоклеточных животных это может быть и только часть тела, например голова) на определенный угол.
Клинотаксисы обнаруживаются и при встрече с твердыми преградами. Вот пример соответствующего поведения инфузории (туфельки). Наткнувшись на твердую преграду (или попав в зону с другими неблагоприятными параметрами среды), туфелька останавливается, изменяется характер биения ресничек, и животное отплывает немного назад. После этого инфузория поворачивается на определенный угол и снова плывет вперед. Это продолжается до тех пор, пока она не проплывет мимо преграды (или не минует неблагоприятную зону).
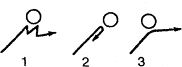
Рис. 27. Зависимость поведения инфузории от угла встречи с преградой (по Розе). Пояснения см. в тексте
Согласно новым данным (В. Розе), инфузории, однако, не всегда ведут себя подобным образом. Клинотаксисы наблюдаются лишь при встрече с преградой под углом в 65–85° (рис. 27, 1). Если же встреча произойдет под прямым углом, инфузория переворачивается через поперечную ось тела и «отскакивает» назад (рис 27, 2). Если же животное встречается с преградой по касательной, оно проплывает мимо, свернув в сторону (рис. 27, 3).
Клинотаксисы хотя встречаются и у более высокоорганизованных животных, относятся к примитивным видам таксисов. Тропотаксисов у простейших нет, поскольку тропотаксисы предполагают наличие симметрично расположенных органов чувств.
В приведенных примерах описывались реакции простейших (в данном случае инфузорий) на температуру и тактильный раздражитель (прикосновение). Речь шла, следовательно, о термо- и тигмотаксисах, в последнем случае – об отрицательном тигмоклинотаксисе, возникающем в ответ на сильное тактильное раздражение (соприкосновение с твердой поверхностью объекта).
Если же, наоборот, туфелька натыкается не на твердое препятствие, а на мягкий объект (например, растительные остатки, фильтровальная бумага), она реагирует иначе: при такой слабой тактильной стимуляции инфузория останавливается и прикладывается к этой поверхности так, чтобы максимальный участок тела соприкасался с поверхностью объекта (положительный тигмотаксис). Аналогичная картина наблюдается и при воздействиях других модальностей на направление движения, т.е. положительный или отрицательный характер реакции зависит от интенсивности раздражения. Как правило, простейшие реагируют на слабые раздражения положительно, на сильные – отрицательно, но в целом простейшим больше свойственно избегать неблагоприятных воздействий, нежели активно искать положительные раздражители.
Возвращаясь к тигмотаксисам, важно отметить, что у инфузорий обнаружены специальные рецепторы тактильной чувствительности – осязательные «волоски», которые особенно выделяются на переднем и заднем концах тела. Эти образования служат не для поиска пищи, а только для тактильного обследования поверхностей объектов, с которыми животное сталкивается. Раздражение этих органелл и приводит в описанном примере к прекращению кинетической реакции.
Особенностью тигмотаксисной реакции является то, что она часто ослабевает, а затем и прекращается после прикасания к объекту максимальной поверхностью тела: приставшая к объекту туфелька в возрастающей мере начинает реагировать на иной раздражитель и все больше отделяется от объекта. Затем, наоборот, вновь возрастает роль тактильного раздражителя и т.д. В результате животное совершает возле объекта ритмичные колебательные движения.
Четко выражена у туфельки и ориентация в вертикальной плоскости, что находит свое выражение в тенденции плыть вверх (отрицательный геотаксис – ориентация по силе земного притяжения). Поскольку у парамеции не были обнаружены специальные органеллы гравитационной чувствительности, было высказано предположение, что содержимое пищеварительных вакуолей действует у нее наподобие статоцистов* высших животных. Обоснованность такого толкования подтверждается тем, что туфелька, проглотившая в опыте металлический порошок, плывет уже не вверх, а вниз, если над ней поместить магнит. В таком случае содержимое вакуоли (металлический порошок) уже давит не на нижнюю ее часть, а, наоборот, на верхнюю, чем, очевидно, и обусловливается переориентация направления движения животного на 180°
* Статоцист – орган чувства равновесия, основная часть которого состоит из замкнутой полости с включенным в нее статолитом – «камешком» (минеральной конкрецией).
Кроме упомянутых таксисные реакции установлены у простейших также в ответ на химические раздражения (хемотаксисы), электрический ток (гальванотаксисы) и др. На свет часть простейших реагируют слабо, у других же эта реакция выражена весьма четко. Так, фототаксисы проявляются у некоторых видов амеб и инфузорий в отрицательной форме, а инфузория Blepharisma снабжена даже светочувствительным пигментом, но, как правило, инфузории не реагируют на изменения освещения. Интересно отметить, что у слабо реагирующих на свет простейших отмечались явления суммации реакции на механические раздражения, если последние сочетались со световыми раздражениями. Такое сочетание может увеличить реакцию в восемь раз по сравнению с реакцией на одно лишь механическое раздражение.
В отличие от инфузорий у многих жгутиковых, особенно у эвглены, положительный фототаксис выражен весьма четко. Биологическое значение этого таксиса не вызывает сомнений, так как аутотрофное питание эвглены требует солнечной энергии. Эвглена плывет к источнику света по спирали, одновременно, как уже упоминалось, вращаясь вокруг собственной оси. Это имеет существенное значение, так как у эвглены, как я у некоторых других простейших, сильно и положительно реагирующих на свет, имеются хорошо развитые фоторецепторы. Это пигментные пятна, иногда снабженные даже отражающими образованиями, позволяющими животному локализовать световые лучи. Продвигаясь к источнику света описанным образом, эвглена поворачивает к нему то «слепую» (спинную) сторону, то «зрячую» (брюшную). И каждый раз, когда последняя (с незаслоненным участком «глазка») оказывается обращенной к источнику света, производится корректировка траектории движения путем поворота на определенный угол в сторону этого источника. Следовательно, движение эвглены к свету определяется положительным фотоклинотаксисом, причем в случае попадания ее под воздействие двух источников света попеременное раздражение фоторецептора то слева, то справа придает движению эвглены внешнее сходство с тропотаксисным поведением двусторонне-симметричных животных, обладающих парными глазами (рис. 28), о чем пойдет речь ниже.
«Глазки» описаны и у других жгутиковых. Например, массивный ярко-красный глазок имеет Collodictyon sparsevacuolata – своеобразный протист с 2–4 жгутиками и амебоидным обликом, способный быстро передвигаться как с помощью жгутиков, так и с помощью ложноножек. Особую сложность фоторецепция достигает у жгутикового Pauchetia (Dinoflagellata), у которого имеются уже аналоги существенных частей глаза многоклеточных животных, пигментное пятно снабжено не только светонепроницаемым экраном (аналог пигментной оболочки), но и све-топроницаемым образованием в форме сферической линзы (аналог хрусталика). Такой «глазок» позволяет не только локализовать световые лучи, но и собирать, в известной степени фокусировать их.
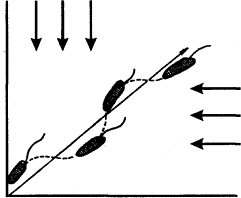
Рис. 28. Траектория передвижения эвглены под воздействием двух одинаково интенсивных источников света (по Будеру)
Пластичность поведения простейших
Как мы видим, и в моторной и в сенсорной сфере поведение достигает у ряда видов простейших известной сложности. Достаточно указать на фобическую реакцию (реакцию испуга) туфельки в вышеописанном примере клинотаксиса: наткнувшись на твердое препятствие (или попав в иную неблагоприятную зону), туфелька резко останавливается и принимает «оборонительное положение», т.е. «съеживается», готовясь пустить в ход ядовитые стрекательные капсулы. Одновременно меняются движения ресничек, происходит тактильное и химическое обследование объекта и т.д. У эвглены фобическая реакция выражается в том, что она, остановившись, начинает производить передним концом тела круговые движения, после чего уплывает в другом направлении (рис. 29).
Ясно, что такая интеграция моторно-сенсорной активности возможна лишь с помощью специальных функциональных структур, аналогичных нервной системе многоклеточных животных. Однако о морфологии этих аналогов еще очень мало известно, и только относительно инфузории удалось с определенной достоверностью доказать существование специальной сетевидной системы проводящих путей, располагающейся в эктоплазме. Очевидно, проведение импульсов осуществляется у простейших и системой градиентов в самой цитоплазме.
|
|
|
Рис. 29. Фобическая реакция эвглены при внезапном изменении освещения (по Джен-нингсу) |
Способность к индивидуальному изменению описанных генетически фиксированных форм поведения путем научения выражена у простейших слабо. Все же способность простейших к научению, хотя бы в элементарных формах, сейчас можно считать доказанной. Если, например, держать парамеции некоторое время в треугольном или квадратном (в сечении) сосуде, то они сохраняют привычный путь передвижения вдоль стенок сосуда и после их перемещения в сосуд округлой формы (рис. 30). Аналогичным образом инфузория, плававшая длительное время (около двух часов) в сосуде, имевшем форму треугольника, придерживается этой траектории затем и в квадратном сосуде большей площади (опыты немецкого ученого Ф.Брамштедта). В опытах советской исследовательницы Н. А.Т ушмаловой инфузории были подвергнуты постоянно действующему ритмичному раздражителю – вибрации. В результате животные постепенно переставали реагировать на него обычным образом (сокращением). Исследовательница усматривает в этом пример элементарных следовых реакций, представляющих собой простую форму кратковременной памяти, формирующейся на этом уровне филогенеза за счет чисто молекулярных взаимодействий.
Подобные опыты ставились и другими экспериментаторами, применявшими различные формы воздействия. Высказывались сомнения, действительно ли в таких случаях можно говорить о каких-то формах научения, ибо сильные воздействия могли оказать хотя бы временное вредное влияние на подопытных животных. К тому же вообще в опытах с простейшими очень трудно учесть все возможные побочные факторы (особенно химические изменения среды).
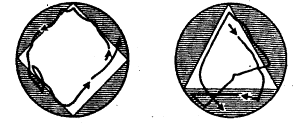
Рис. 30. Привыкание у парамеции (по Брамштедту). Пояснения см. в тексте
Все же, очевидно, мы имеем здесь дело с элементарной формой научения – привыканием. Привыкание (к измененным внешним условиям), как мы уже знаем, играет немаловажную роль и в поведении высших животных, но имеет качественно иной характер уже хотя бы потому, что строится у них не только на ощущениях, но и на восприятиях. При элементарной же сенсорной психике животное может привыкать лишь к воздействию отдельных раздражителей (или их совокупностей), являющихся воплощением отдельных свойств или качеств компонентов окружающей среды. Именно это и имело место в приведенных примерах, когда наблюдалась модификация видотипичного поведения: врожденные реакции на определенные раздражители постепенно элиминируются, если при многократном повторении не наступит биологически значимый эффект. Так, в опытах Тушмаловой фобическая реакция у инфузорий со временем сводилась на нет при повторной встрече с «безобидным» раздражителем.
Привыкание необходимо отличать от утомления, выражающегося в явлениях «истощения» животного. В опытах Тушмаловой это выражалось в неспособности инфузории к дальнейшему реагированию, если она в течение 2–3 часов подвергалась очень сильным раздражениям. Утомление связано с перерасходом энергетических ресурсов, привыкание же представляет собой активную приспособительную реакцию, значение которой состоит в экономии этих ресурсов, в предотвращении траты энергии на бесполезные для животного движения.
Как форма научения привыкание характеризует низший уровень элементарной сенсорной психики, хотя и не теряет своего значения на всех ступенях развития психики, как это было показано при ознакомлении с онтогенезом поведения животных (см. ч. II). Однако у высших представителей простейших, возможно, уже существуют и зачатки ассоциативного научения, вообще характерного для более высоких уровней психического развития.
Об ассоциативном научении можно говорить в тех случаях, когда устанавливается временная связь между биологически значимым и «нейтральным», точнее, биологически маловалентным раздражителем. Именно такую связь Брамштедту удалось выработать у туфельки, которая, как уже отмечалось, не реагирует заметным образом на изменение освещения, но весьма чувствительна к температурным изменениям. Если в эксперименте затенить одну половину капли воды, в которой плывут парамеции, и при этом подогреть освещенную часть капли, то вскоре инфузории соберутся в холодной темной части, но останутся там (в течение 15 минут) и после того, как температура в обеих частях капли будет уравновешена. Правда, эти опыты Брамштедта подвергались серьезной критике, так как при подогреве меняется и химизм воды (меняется растворимость содержащихся в ней газов), что не может не влиять на поведение инфузории (опыты У.Грабовского).
Рис. 31. Образование у парамеции временной связи между электрическим раздражением и светом (по Зесту). Пояснения см. в тексте
Вместе с тем аналогичный результат может быть получен и путем наказания животных при заплыве в освещенную зону ударами электрического тока (рис. 31). Эффект избегания сохранялся в этом случае в течение 20 минут (опыты Г.Зеста). Однако обратный результат, т.е. избегание темноты и заплыв в освещенную зону, Зесту получить не удалось. Это обстоятельство, однако, является результатом методической ошибки: инфузориям свойствен гальванотаксис, выражающийся в их устремлении к катоду. В упомянутых же опытах Зеста катод всегда находился на темной половине и гальванотаксис оказывался сильнее электрического раздражения, почему инфузории и направлялись только в темноту.
В более тщательно поставленных опытах польскому ученому С.Вавржинчику удалось преодолеть эти методические недостатки и с успехом научить инфузории избегать темный участок стеклянной трубки, где их раздражали электрическим током. Постепенно туфельки все чаще оставались в освещенной зоне и поворачивали на границе с затемненной зоной даже при перемещении этой границы вдоль трубки еще до получения электрического удара. Наконец, они оставались в освещенной зоне в четыре раза дольше, чем в темной, даже тогда, когда раздражения током совсем прекратились. Эта реакция сохранялась даже в течение 50 минут, что является достаточно большим сроком для простейших. Четкие положительные результаты этот исследователь получил и в других экспериментах.
Правда, Я.Дембовский, повторивший эти опыты с некоторыми изменениями, пришел к выводу, что, хотя и не исключена возможность выработки у простейших обусловленных реакций, эти опыты с туфельками все-таки не решают до конца сути вопроса, ибо существуют несравненно более сильные факторы, особенно химические, которые маскируют реакции на свет. Ввиду отсутствия вполне убедительных экспериментальных данных Дембовский оставил вопрос о наличии у простейших обусловленных реакций открытым.
Вместе с тем и у других (кроме туфельки) инфузорий, в частности у стентора и сувойки, удалось после 140–160 сочетаний выработать сократительные реакции в ответ на световое раздражение.
На инфузориях проводились и другие опыты с применением разных методических приемов. В частности, использовались капилляры, иногда с загнутым концом, в которые сажались парамеции, после чего измерялось время, необходимое им для высвобождения. В новейших исследованиях, проведенных таким способом, получались вполне положительные результаты: при каждом повторении опыта время выхода парамеции из трубочки заметно уменьшалось, что и признается доказательством их способности к ассоциативному научению.
Однако американские ученые Ф.Б.Эпплуайт и Ф.Т.Гарднер, недавно повторившие эти опыты, считают такой вывод необоснованным. Эти ученые засасывали последовательно в один и тот же капилляр разные, еще ни разу не побывавшие в опыте туфельки и установили, что время выхода этих животных из капилляра также постепенно уменьшается. Но этого не случалось, если капилляр до употребления тщательно промывался и кипятился в деионизированной воде. Использование обработанных таким образом капилляров для повторного проведения опыта с одной и той же туфелькой не давало практически никакого эффекта научения. Отсюда экспериментаторы вывели заключение, что уменьшение времени выхода из капилляра является у парамеции не результатом научения, а реакцией на загрязнение внутренней поверхности трубки продуктами обмена веществ, которое увеличивается от опыта к опыту и от которого животные пытаются все скорее уйти.
С другой стороны, в аналогичных исследованиях других ученых учитывались подобные возможные изменения среды, и было установлено, что время выхода инфузорий из трубки сокращается лишь в первых опытах, потом же оно остается постоянным (среда в трубке должна, однако, продолжать изменяться).
Таким образом, вопрос о наличии у инфузорий (а тем более у простейших вообще) ассоциативного научения еще нельзя считать решенным. Все же, очевидно, такое научение существует у них в зачаточном виде.
Общая характеристика психической активности
Как мы могли убедиться, на низшем уровне элементарной сенсорной психики поведение животных выступает в достаточно разнообразных формах, но все же мы имеем здесь дело лишь с примитивными проявлениями психической активности. О такой активности, о психике мы можем здесь говорить потому, что простейшие активно реагируют на изменения в окружающей их среде, причем реагируют на биологически непосредственно не значимые свойства компонентов среды как на сигналы о появлении жизненно важных условий среды. Другими словами, простейшим свойственна элементарная форма психического отражения – ощущение, т.е. чувствительность в собственном смысле слова. Но там, как мы уже знаем, где появляется способность к ощущению, начинается психика.
Уже говорилось и о том, что даже низший уровень психического отражения не является низшим уровнем отражения вообще, существующего в живой природе: растениям присуще допсихическое отражение, при котором имеют место лишь процессы раздражимости. Однако элементы такого допсихического отражения встречаются и у простейших.
Большой интерес представляют в этом отношении, как и вообще для понимания условий зарождения психического отражения, реакции простейших на температуру. Здесь непосредственно необходимая для поддержания жизни энергия еще тождественна опосредующей энергии, сигнализирующей о наличии жизненно необходимого компонента среды.
Еще в начале нашего века М.Мендельсон установил, что у инфузорий реакции на изменения температуры становятся все более дифференцированными по мере приближения к некому термическому оптимуму. Так, у Paramecium aurelia такой оптимум находится в пределах +24 ... +28° температуры воды. При +6 ... +15° инфузория реагирует на разности температуры от 0,06 до 0,08°, при +20 ... +24° же – на разности от 0,02 до 0,005°. Но по современным данным этот вид хорошо переносит и низкие температуры и даже замораживание до -30° и более.
Вместе с тем у простейших специфические терморецепторы, т.е. специальные образования, служащие для восприятия изменений температуры среды, не обнаружены. (Достоверно такие рецепторы неизвестны и для других беспозвоночных.) В свое время Г.Дженнингс полагал, что у инфузории (туфельки) передний конец тела особенно чувствителен в отношении температурных раздражений. Однако впоследствии О.Кёлер показал, что у разрезанной на две части туфельки передний и задний концы тела ведут себя в этом отношении в принципе так же, как целое животное. Отсюда напрашивается вывод, что реакция на изменения температуры является у простейших свойством всей протоплазмы. Возможно, что реакции простейших на термические раздражения еще сходны с биохимическими реакциями типа ферментативных процессов.
Очевидно, все это и соответствует тому обстоятельству, что для организма простейшего тепловая энергия еще не расчленяется на источник жизнедеятельности и сигнал о наличии такого источника. В связи с этим отсутствуют и специальные терморецепторы. Мы имеем, следовательно, здесь пример «сосуществования» у простейших допсихического и психического отражения.
Интересно, что отсутствие терморецепторов не делает реакции простейших менее выразительными, чем их реакции на другие внешние агенты. Как было показано, эти реакции проявляются в таксисах (в данном случае термотаксисах) и кинезах.
Еще явственнее «сосуществование» допсихического и психического отражения выступает у низших представителей типа. У эвглены оно обусловлено и наличием наряду с животным, растительного, аутотрофного типа питания, почему эвглена в одинаковой степени относится и к растениям и к животным.
С другой стороны, мы знаем, что качества психического отражения определяются тем, насколько развиты способности к движению, пространственно-временной ориентации и к изменению врожденного поведения. У простейших мы встречаем разнообразные формы передвижения в водной среде, но только на самом примитивном уровне инстинктивного поведения – кинезов. Ориентация поведения осуществляется только на основе ощущений и ограничена элементарными формами таксисов, позволяющими животному избегать неблагоприятные внешние условия. За отдельными исключениями, активность простейших находится в целом как бы под отрицательным знаком, ибо эти животные попадают в сферу действия положительных раздражителей, уходя от отрицательных.
Это значит, что поисковая фаза инстинктивного поведения (кинезов) в указанном отношении еще крайне недоразвита. К тому же она явно лишена сложной, многоэтапной структуры. Возможно, что во многих случаях эта фаза вообще отсутствует. Во всем этом проявляется не только исключительная примитивность инстинктивного поведения на данном уровне, но и предельная скудность содержания психического отражения. Ведь это содержание наполняется прежде всего активным поиском и оценкой раздражителей на первых этапах поисковой фазы.
Как уже отмечалось, в отдельных случаях у простейших встречаются и положительные элементы пространственной ориентации. К уже упомянутым примерам положительных таксисов можно добавить, что амеба в состоянии находить пищевой объект на расстоянии до 20– 30 микрон. Зачатки активного поиска жертвы существуют, очевидно, и у хищных инфузорий. Однако во всех этих случаях положительные таксисные реакции еще не носят характера подлинного поискового поведения, поэтому эти исключения не меняют общую оценку поведения простейших, а тем более характеристику низшего уровня элементарной сенсорной психики в целом: дистантно на этом уровне распознаются преимущественно отрицательные компоненты среды; биологически «нейтральные» же признаки положительных компонентов, как правило, еще не воспринимаются на расстоянии как сигнальные, т.е. попросту еще не существуют для животного как таковые. Таким образом, психическое отражение выполняет на самом низком уровне своего развития в основном сторожевую функцию и отличается поэтому характерной «однобокостью»: сопутствующие биологически не значимые свойства компонентов среды дистантно ощущаются животными как сигналы появления таких компонентов лишь в том случае, если эти компоненты являются для животных вредными.
Что касается, наконец, пластичности поведения простейших, то и здесь простейшие обладают лишь самыми элементарными возможностями. Это вполне закономерно: элементарному инстинктивному поведению может соответствовать лишь элементарное научение. Последнее, как мы видели, представлено наиболее примитивной формой – привыканием, и лишь в отдельных случаях, может быть, встречаются зачатки ассоциативного научения.
Конечно, при всей своей примитивности поведение простейших является все же достаточно сложным и гибким, во всяком случае в тех пределах, которые необходимы для жизни в своеобразных условиях микромира. Эти условия отличаются рядом специфических особенностей, и этот мир нельзя себе представить как просто во много раз уменьшенный макромир. В частности, среда микромира является менее стабильной, чем среда макромира, что проявляется, например, в периодическом высыхании маленьких водоемов. С другой стороны, непродолжительность жизни микроорганизмов как отдельных особей (частая смена поколений) и относительное однообразие этого микромира делают излишним развитие более сложных форм накопления индивидуального опыта. В этой микросреде нет столь сложных и разнообразных условий, к которым можно приспособиться только путем научения. В таких условиях пластичность самого строения простейших, легкость образования новых морфологических структур в достаточной мере обеспечивают приспособляемость этих животных к условиям существования. Можно, очевидно, сказать, что пластичность поведения здесь еще не превзошла пластичность строения организма.
Как уже отмечалось, простейшие не являются однородной группой животных, и различия между их разными формами очень велики. Высшие представители этого типа во многих отношениях развивались в своеобразных формах неклеточного строения параллельно низшим многоклеточным беспозвоночным животным. В результате высокоразвитые простейшие проявляют подчас даже более сложное поведение, чем некоторые многоклеточные беспозвоночные, также стоящие на низшем уровне элементарной сенсорной психики. Это тоже одна из причин, почему мы описали этот уровень психического развития на примере только простейших. Здесь наглядно выступает уже отмеченная общая закономерность: психологическая классификация не вполне совпадает с зоологической, так как некоторые представители одной и той же таксономической категории могут еще находиться на более низком психическом уровне, другие – уже на более высоком. Именно последнее имеет место у высших представителей типа простейших, которых в этом отношении можно было бы рассматривать как исключения. Однако, в сущности, это не так, ибо здесь проявляется и другая закономерность эволюции психики, а именно: элементы более высокого уровня психического развития всегда зарождаются в недрах предшествующего, более низкого уровня. В данном случае, например, примитивные формы ассоциативного научения, вообще характерные для более высокого уровня элементарной сенсорной психики, встречаются в зачатке уже у некоторых видов, относящихся к типу, который в целом стоит на низшем уровне элементарной сенсорной психики, где типичной формой индивидуально-изменчивого поведения является привыкание.
Высший уровень развития элементарной сенсорной психики
Высшего уровня элементарной сенсорной психики достигло большое число многоклеточных беспозвоночных. Однако, как отмечалось, часть низших многоклеточных беспозвоночных находится в основном на том же уровне психического развития, что и многие простейшие. Это относится прежде всего к большинству кишечнополостных и к низшим червям, а тем более к губкам, которые во многом еще напоминают колониальные формы одноклеточных (жгутиковых). Неподвижный, сидячий образ жизни взрослых губок привел даже к редукции их внешней активности, поведения (при полном отсутствии нервной системы и органов чувств). Но даже у самых примитивных представителей многоклеточных животных создались принципиально новые условия поведения в результате появления качественно новых структурных категорий – тканей, органов, систем органов. Это и обусловило возникновение специальной системы координации деятельности этих многоклеточных образований и усложнившегося взаимодействия организма со средой – нервной системы.
К низшим многоклеточным беспозвоночным относятся помимо уже упомянутых еще иглокожие, высшие (кольчатые) черви, отчасти моллюски и др.
Мы рассмотрим в дальнейшем в качестве примера кольчатых червей, у которых в полной мере выражены признаки поведения, характерные для высшего уровня элементарной сенсорной психики. К кольчатым червям относятся живущие в морях многощетинковые черви (полихеты), малощетинковые черви (наиболее известный представитель – дождевой червь) и пиявки. Характерным признаком строения кольчецов является внешняя и внутренняя метамерия: тело состоит из нескольких большей частью идентичных сегментов, каждый из которых содержит «комплект» внутренних органов, в частности пару симметрично расположенных ганглиев с нервными коммисурами. В результате нервная система кольчатых червей имеет вид «нервной лестницы».
Нервная система
Как известно, нервная система впервые появляется у низших многоклеточных беспозвоночных. Возникновение нервной системы – важнейшая веха в эволюции животного мира, и в этом отношении даже примитивные многоклеточные беспозвоночные качественно отличаются от простейших. Важным моментом здесь является уже резкое ускорение проводимости возбуждения в нервной ткани: упротоплазме скорость проведения возбуждения не превышает 1–2 микрон в секунду, но даже в наиболее примитивной нервной системе, состоящей из нервных клеток, она составляет 0,5 метра в секунду!
Нервная система существует у низших многоклеточных в весьма разнообразных формах: сетчатой (например, у гидры), кольцевой (медузы), радиальной (морские звезды) и билатеральной. Билатеральная форма представлена у низших (бескишечных) плоских червей и примитивных моллюсков (хитон) еще только сетью, располагающейся вблизи поверхности тела, но выделяются более мощным развитием несколько продольных тяжей. По мере своего прогрессивного развития нервная система погружается под мышечную ткань, продольные тяжи становятся более выраженными, особенно на брюшной стороне тела. Одновременно все большее значение приобретает передний конец тела, появляется голова (процесс цефализации), а вместе с ней и головной мозг – скопление и уплотнение нервных элементов в переднем конце. Наконец, у высших червей центральная нервная система уже вполне приобретает типичное строение «нервной лестницы», при котором головной мозг располагается над пищеварительным трактом и соединен двумя симметричными коммисурами («окологлоточное кольцо») с расположенными на брюшной стороне подглоточными ганглиями и далее с парными брюшными нервными стволами. Существенными элементами являются здесь ганглии, поэтому говорят и о ганглионарной нервной системе, или о «ганглионарной лестнице». У некоторых представителей данной группы животных (например, пиявок) нервные стволы сближаются настолько, что получается «нервная цепочка».
От ганглиев отходят мощные проводящие волокна, которые и составляют нервные стволы. В гигантских волокнах нервные импульсы проводятся значительно быстрее благодаря их большому диаметру и малому числу синаптических связей (мест соприкосновения аксонов одних нервных клеток с дендритами и клеточными телами других клеток). Что же касается головных ганглиев, т.е. мозга, то они больше развиты у более подвижных животных, обладающих и наиболее развитыми рецепторными системами.
Зарождение и эволюция нервной системы обусловлены необходимостью координации разнокачественных функциональных единиц многоклеточного организма, согласования процессов, происходящих в разных частях его при взаимодействии с внешней средой, обеспечения деятельности сложно устроенного организма как единой целостной системы. Только координирующий и организующий центр, каким является центральная нервная система, может обеспечить гибкость и изменчивость реакции организма в условиях многоклеточной организации.
Огромное значение имел в этом отношении и процесс цефализапии, т.е. обособления головного конца организма и сопряженного с ним появления головного мозга. Только при наличии головного мозга возможно подлинно централизованное «кодирование» поступающих с периферии сигналов и формирование целостных «программ» врожденного поведения, не говоря уже о высокой степени координации всей внешней активности животного.
Разумеется, уровень психического развития зависит не только от строения нервной системы. Так, например, близкие к кольчатым червям коловратки также обладают, как и те, билатеральной нервной системой и мозгом, а также специализированными сенсорными и моторными нервами. Однако, мало отличаясь от инфузории размером, внешним видом и образом жизни, коловратки очень напоминают последних также поведением и не обнаруживают более высоких психических способностей, чем инфузории. Это опять показывает, что ведущим для развития психической деятельности является не общее строение, а конкретные условия жизнедеятельности животного, характер его взаимоотношений и взаимодействий с окружающей средой. Одновременно этот пример еще раз демонстрирует, с какой осторожностью надо подходить к оценке «высших» и «низших» признаков при сравнении организмов, занимающих различное филогенетическое положение, в частности при сопоставлении простейших и многоклеточных беспозвоночных.
Движения
Кольчатые черви обитают в морях и пресноводных водоемах, но некоторые ведут и наземный образ жизни, передвигаясь ползком по субстрату или роясь в рыхлом грунте. Морские черви отчасти пассивно носятся течениями воды как составная часть планктона, но основная масса ведет придонный образ жизни в прибрежных зонах, где селится среди колоний других морских организмов или в расщелинах скал. Многие виды живут временно или постоянно в трубках, которые в первом случае периодически покидаются их обитателями, а затем вновь разыскиваются. Хищные виды отправляются из этих убежищ регулярно на «охоту». Трубки строятся из песчинок и других мелких частиц, которые скрепляются выделениями особых желез, чем достигается большая прочность построек. Неподвижно сидящие в трубках животные ловят свою добычу (мелкие организмы), подгоняя к себе и процеживая воду с помощью венчика щупалец, который высовывается из трубки, или же прогоняя сквозь нее поток воды (в этом случае трубка открыта на обоих концах).
В противоположность сидячим формам свободноживущие черви активно разыскивают свою пищу, передвигаясь по морскому дну: хищные виды нападают на других червей, моллюсков, ракообразных и иных сравнительно крупных животных, которых хватают челюстями и проглатывают; растительноядные отрывают челюстями куски водорослей; другие черви (их большинство) ползают и роются в придонном иле, проглатывают его вместе с органическими остатками или собирают с поверхности дна мелкие живые и мертвые организмы.
Малощетинковые черви ползают и роются в мягком грунте или придонном иле, некоторые виды способны плавать. Во влажных тропических лесах некоторые малощетинковые кольчецы вползают даже на деревья. Основная масса малощетинковых червей питается детритом, всасывая слизистый ил или прогрызаясь сквозь почву. Но существуют и виды, поедающие мелкие организмы с поверхности грунта, процеживающие воду или отгрызающие куски растений. Несколько видов ведут хищный образ жизни и захватывают мелких водных животных, резко открывая ротовое отверстие. В результате добыча всасывается с потоком воды.
Пиявки хорошо плавают, производя туловищем волнообразные движения, ползают, роют ходы в мягком грунте, некоторые передвигаются по суше. Помимо кровососущих существуют также пиявки, которые нападают на водных беспозвоночных и проглатывают их целиком. Наземные пиявки подстерегают свои жертвы на суше (млекопитающих и людей), в траве или на ветках деревьев и кустарников (во влажных тропических лесах). Эти пиявки могут довольно быстро двигаться. В передвижении наземных пиявок по субстрату большую роль играют присоски: животное вытягивает сперва туловище, затем присасывается к субстрату головной присоской, притягивает к ней задний конец туловища (с одновременным сокращением последнего), присасывается задней присоской и т.д.
Итак, двигательная активность кольчатых червей как при локомоции, так и при добывании пищи отличается большим многообразием и достаточной сложностью. Обеспечивается это сильно развитой мускулатурой, представленной прежде всего так называемым кожно-мышечным мешком. Он состоит из двух слоев: внешнего (подкожного), состоящего из кольцевых волокон, и внутреннего, состоящего из мощных продольных мышц. Последние простираются, несмотря на сегментацию, от переднего до заднего конца туловища. Ритмичные сокращения продольной и кольцевой мускулатуры кожно-мышечного мешка обеспечивают локомоцию: червь ползет, вытягивая и сокращая, расширяя и сужая отдельные части своего тела. Так, у дождевого червя вытягивается (и сужается) передняя часть тела, затем то же самое происходит последовательно со следующими сегментами. В результате по телу червя пробегают «волны» сокращений и расслаблений мускулатуры.
У кольчатых червей впервые в эволюции животного мира появляются подлинные парные конечности: кольчецы (помимо пиявок) носят на каждом сегменте по паре выростов, служащих органами передвижения (кроме головного конца, где они служат ротовыми органами) и получивших название параподий. Параподии снабжены специальными мышцами, двигающими их вперед или назад. Зачастую параподий имеют ветвистое строение. Каждая ветвь снабжена опорной щетинкой и, кроме того, венчиком из щетинок, имеющих у разных видов различную форму. От параподий отходят и щупальцевидные органы тактильной и химической чувствительности. Особенно длинными и многочисленными последние являются на головном конце, где на спинной стороне располагаются глаза (одна или две пары), а в ротовой полости или на особом (выпячиваемом) хоботке – челюсти. В захвате пищевых объектов могут участвовать и нитевидные щупальца на головном конце червя.
У ряда многошетинковых и всех малощетинковых червей параподии редуцированы (отсюда и название последних), остались лишь посегментно расположенные пучки щетинок. Так, у дождевого червя на каждом сегменте находится по четыре пары очень коротких, неразличимых невооруженным глазом щетинок, которые, однако, наподобие параподии служат для передвижения животного: являясь достаточно крепкими подвижными рычагами, они обеспечивают вместе с сокращениями кожно-мышечного мешка поступательное движение червя. С другой стороны, растопыривая свои щетинки и упираясь ими в грунт, дождевой червь настолько прочно фиксирует свое тело в земле, что практически невозможно вытащить его оттуда в неповрежденном виде. У некоторых других малощетинковых червей щетинки развиты значительно сильнее и представлены в большем количестве.
Органы чувств и сенсорные способности
Большой интерес для познания психической деятельности низших многоклеточных беспозвоночных представляет устройство и функционирование их органов чувств, представленных также весьма различными образованиями в соответствии с общим уровнем организации животного.
У наиболее низкоорганизованных представителей беспозвоночных органы чувств еще очень слабо дифференцированы как в морфологическом, так и в функциональном отношении. У этих животных трудно выделить органы осязания, химической чувствительности и т.д. Очевидно, первичные органы чувств вообще были плюромодальными, т.е. они обладали лишь общей, присущей всей живой материи чувствительностью, но в повышенной степени. Существование таких плюромодальных чувствительных клеток является весьма вероятной гипотезой, но в настоящее время такие рецепторные клетки, очевидно, уже не существуют. Специализация таких клеток по отдельным видам энергии привела к появлению унимодальных рецепторных образований, которые, как правило, реагируют лишь на один специфический вид энергии. Так появились термо-, хемо-, механо-, фото- и другие рецепторы.
Согласно этой гипотезе, все органы чувств многоклеточных животных развились из органов осязания – наименее дифференцированных рецепторов. В наиболее элементарных случаях осязательная функция присуща всем клеткам поверхности тела. Но уже у кишечнополостных появляются специальные осязательные клетки, которые, скапливаясь в определенных местах, образуют подлинные органы осязания. Это вытянутые цилиндрические или веретеновидные клетки, несущие на конце неподвижный чувствительный волосок или пучок волосков. Однако эти органы часто выполняют и обонятельную функцию. Особенностью низших многоклеточных беспозвоночных является то, что во многих (хотя и не во всех) случаях эти две рецепторные функции слиты и не поддаются морфологическому разграничению.
В этом нетрудно усмотреть остатки первичной плюромодальности.
С другой стороны, органы зрения относятся к наиболее сложным по строению и функционированию. Иногда органы чувств низшего порядка превращаются в органы чувств высшего порядка (например, у пиявок некоторые из органов осязания – так называемые «сенсиллы» – превращаются в глаза). Известный советский зоолог, специалист по сравнительной анатомии беспозвоночных B-А.Догель говорил в таких случаях о «повышении органа в ранге». Однако в процессе филогенеза, как отмечает Догель, нередко имело место и обратное явление; в других случаях какие-то рецепторы беспозвоночных исчезали, чтобы потом вновь появиться в несколько измененной форме. Это непостоянство и легкость перестройки привели к тому, что у близкородственных беспозвоночных однотипные рецепторы, в частности органы оптической чувствительности, подчас бывают совершенно различными по строению и функциям.
Если взять, к примеру, кишечнополостных, то гидра четко реагирует на свет, хотя специальных органов зрения у нее нет. Она воспринимает свет всей поверхностью тела. Положительный фототаксис гидры выражается в том, что животное производит в освещенной сфере круговые или маятникообразные колебательные движения и в конце концов занимает положение в сторону источника света или даже направляется (ползет) к нему. Свободноживущие же представители кишечнополостных – медузы – обладают уже специальными многоклеточными органами светочувствительности. В простейшем случае эти органы представлены так называемыми глазными пятнами, которые находятся среди обыкновенных эпителиальных клеток, и даже нечетко отграничены от них. Более дифференцированным рецептором является глазная ямка (рис. 32). Однако чаще у медуз встречаются уже настоящие глаза, причем наиболее сложно устроенные из них представляют собой погруженные под слоем эпителиальных клеток глазные пузыри приблизительно шарообразной формы. Эпителий над глазным пузырем утончен и представляет собой прозрачную роговицу. Дно и стенки пузыря состоят из двух типов клеток: ретинальных и пигментных, причем ретинальные клетки снабжены чувствительными палочками. В полости глазного пузыря находится стекловидное тело – студенистая масса, защищающая ретину от механических повреждений. Иногда встречаются даже хрусталик и радужка, и тогда налицо все основные компоненты глаза высших животных (нет, однако, глазодвигательных мышц и систем фокусировки).
Рис. 32. Развитие глаза беспозвоночных (на примере гидромедуз). Слева вверху – глазное пятно; слева внизу – глазная ямка; справа – глазной пузырь. 1 – зрительная клетка; 2 – пигментная клетка; 3 – зрительный нерв; 4 – эпителий; 5 – хрусталик; 6 – светочувствительные палочки; 7–стекловидное тело (по Бючли)
Учитывая, что глазное пятно является исходной формой вообще всех органов зрения, можно, следовательно, в ряду медуз проследить путь усложнения структуры от самого примитивного органа светочувствительности до сложного, высокодифференцированного глаза.
Очень разнообразны по своему строению и глаза червей, как и других низших многоклеточных беспозвоночных; в принципе к ним относится то же, что говорилось в отношении глаз медуз. В соответствии с многообразием движений кольчатых червей, разнообразием способов добывания пищи и других моментов жизнедеятельности находится и уровень развития сенсорной сферы этих животных. Это, правда, не означает, что у кольчецов имеются рецепторы для всех видов энергии, воздействующих на них, или даже что для всех форм чувствительности имеются специальные органы чувств. Так, например, у кольчатых червей встречаются сложно устроенные глаза, снабженные даже хрусталиками. Имеются весьма сложные глаза и у некоторых планарий и улиток. По их обладатели, насколько известно, неспособны к зрительному восприятию предметов. За исключением, может быть, некоторых улиток, у всех этих животных фотоскопические глаза, позволяющие отличать свет от тьмы и направление, откуда световые лучи падают на животное, а также перемещение светотеней в непосредственной близости от животного. Светочувствительность может при этом быть очень высокой, например, гребешок, двустворчатый моллюск с несколькими десятками глаз, закрывает створки раковины уже при уменьшении интенсивности освещения на 0,3%. (Для сравнения можно указать, что человек воспринимает уменьшение освещения лишь не менее чем на 1%).
Большой интерес представляют активно плавающие многощетинковые кольчатые черви из семейства Alciopidae, ведущие хищный образ жизни. У этих полихет глаз не только отличается исключительно сложным строением и величиной, но и снабжен аккомодационным устройством в виде специальных сократительных волокон, способных передвигать хрусталик и тем самым менять фокусное расстояние. Это единственный известный случай среди низших беспозвоночных: аккомодация глаза встречается только у головоногих моллюсков и позвоночных. Возможно, у этих червей в какой-то степени уже существует предметное зрение, что было бы исключением, подтверждающим общее правило. Это относится и к свободно плавающим хищным моллюскам Heteropoda, которые тоже обладают весьма сложно устроенными глазами с приспособлением, заменяющим аккомодацию.
Что же касается дождевого червя, то здесь обнаруживается чрезвычайно интересный факт: у него нет не только сложно устроенных, но и вообще никаких специальных органов светочувствительности. Вместе с тем ему свойствен четкий отрицательный фототаксис. Функцию светоощущения выполняют рассеянные в коже светочувствительные клетки. Это пример кожной светочувствительности низших многоклеточных беспозвоночных. Кожная светочувствительность наблюдается и у многих моллюсков, причем у двустворчатых это нередко единственная форма фоторецепции. Эти моллюски реагируют как на освещение, так и на затемнение чаще всего одинаковым образом – втягиванием выступающих из раковин частей тела или запиранием раковины. Многие улитки реагируют на внезапное затемнение сокращением «ноги», причем эта реакция сохраняется и после экстирпации глаз, что опять-таки указывает на наличие кожной светочувствительности.
Реакция дождевого червя на условия освещения состоит в том, что он уползает в зону большего затемнения. Если же внезапно уменьшить интенсивность освещения, то червь реагирует на это движениями бегства; в естественных условиях он вползает в почву. Такая же реакция следует за внезапным освещением. Если же осветить лишь определенный участок тела дождевого червя, то время этой реакции сокращается пропорционально размеру освещаемой площади поверхности тела – в наибольшей мере червь реагирует при освещении всей его поверхности. Следовательно, реакция определяется градиентом раздражения освещенных и неосвещенных частей тела. Аналогично реагируют на свет и другие кольчатые черви.
Кольчатые черви реагируют также на прикосновения, химические и термические раздражения, силу тяжести, электрические раздражения, течение воды, а наземные формы (дождевые черви) – на влажность среды. Однако принципиальных отличий от реакции на свет нет: все эти реакции находятся на одном уровне и характеризуются тем, что являются ответами на отдельные раздражители, на отдельные признаки, качества предметов, но не на сами предметы как таковые. Так, например, много- и малощетинковые черви проявляют четко выраженные таксисные реакции на тактильные раздражения. Преобладают отрицательные реакции, но в ряде случаев наблюдаются и положительные тигмотаксисы: прикосновения нервных сегментов животного к субстрату влекут за собой прижимание к нему всем телом, что имеет, конечно, большое значение при роющем образе жизни или жизни в трубках. Интересно, что у дождевого червя соответствующие рецепторные образования представлены лишь отдельными чувствительными клетками, разбросанными по всему телу, но более густо располагающимися на его переднем конце. У многощетинковых же червей органами осязания часто являются щупальца или щетинки.
Хорошо развита и химическая чувствительность, причем в большинстве случаев наблюдается отрицательный хемотаксис. При большой интенсивности химического воздействия черви всегда реагируют отрицательно. С другой стороны, дождевой червь, например, способен по химическим признакам выбирать разные виды листьев, что свидетельствует о специализации в сенсорной сфере. У ряда кольчецов обнаружены расположенные около ротового отверстия органы химической чувствительности в виде ямок. Особенно это относится к пларающим видам, у которых имеется пара таких ямок, выстланных мерцательным эпителием.
Таксисы
Как и на более низких уровнях эволюционного развития, пространственная ориентация совершается на высшем уровне элементарной сенсорной психики, преимущественно на основе примитивных таксисов. Но с усложнением жизнедеятельности организмов возрастают и требования к локализации биологически значимых компонентов среды по их биологически незначимым признакам. Возникает необходимость более сложного таксисного поведения, позволяющего животному достаточно четко и дифференцированно ощущать и реагировать. Именно этим и отличаются животные, находящиеся на рассматриваемом здесь уровне развития психики: благодаря симметричному расположению органов чувств у них обнаруживаются наряду с кинезами и элементарными таксисами и некоторые высшие формы таксисного поведения.
Немецкий ученый А.Кюн выделил следующие категории высших таксисов, которые, правда, вполне развиты лишь у высших животных: тропотаксисы – движение ориентируется по равнодействующей, образуемой в результате выравнивания интенсивности возбуждения в симметрично расположенных рецепторах; телотаксисы – выбор и фиксация одного источника раздражения и направление движения к этому источнику («цели»); менотаксисы – при несимметричном раздражении в симметрично расположенных рецепторах движение производится под углом к источнику раздражения. Менотаксисы играют ведущую роль в сохранении животным константного положения в пространстве.
Кроме того, у высших животных с развитой памятью встречается еще одна форма таксисов – мнемотаксисы, при которых основную роль играет индивидуальное запоминание ориентиров, что особенно важно для территориального поведения, о чем еще пойдет речь.
В поведении кольчецов тропо- и телотаксисы чаще всего проявляются совместно: если животное подвергнуть одновременному воздействию двух источников энергии (например, света) одинаковой силы, то при положительном таксисе оно будет первоначально двигаться по направлению равнодействующей, т.е. линии, ведущей к середине расстояния между источниками (тропотаксис), но затем, как правило, еще не дойдя до этой точки, оно повернет и направится к одному из источников энергии (телотаксис).
Наиболее четко тропо- и телотаксисы проявляются у активно двигающихся кольчатых червей, прежде всего у хищников. Последние не только обнаруживают присутствие жертвы по исходящим от нее физическим и химическим стимулам, но и направляются к ней, ориентируясь на основе таких таксисов. Наземные пиявки, например, способны точно локализовать местонахождение жертвы и с удивительной быстротой направляются к ней, ориентируясь по сотрясению субстрата, а затем, на более близком расстоянии, и по исходящему от жертвы (крупных млекопитающих) теплу и запаху. В результате хищник обнаруживает ее по соответствующим градиентам с помощью сочетания положительных вибрационных, термических и химических телотаксисов.
При менотаксисах животное, как говорилось, двигается под углом к линии воздействия, исходящей от источника энергии. Типичный пример – ориентация мигрирующих животных по солнечному (или другому астрономическому) «компасу», что, конечно, особенно характерно для перелетных птиц. У низших многоклеточных беспозвоночных менотаксисы обнаружены у некоторых улиток. Если этих животных поместить на освещаемый сбоку и медленно вращающийся диск, то они, двигаясь против направления вращения, придерживаются определенного угла по отношению к источнику света. В этих случаях, следовательно, можно уже говорить об ориентации животного по световому компасу, которая играет исключительную роль в жизни высших животных.
Зачатки высших форм поведения
Многощетинковые кольчатые черви относятся к наиболее развитым в психическом отношении низшим беспозвоночным. Их поведение отличается подчас большой сложностью и представляет особый интерес в том смысле, что содержит ряд элементов психической деятельности, присущей более высокоорганизованным животным. В отличие от основной массы низших беспозвоночных у полихет наблюдаются некоторые существенные усложнения видотипичного поведения, отчасти уже выходящие за рамки типичной элементарной сенсорной психики.
Так, в некоторых случаях у этих морских червей встречаются действия, которые уже можно назвать конструктивными, поскольку животные активно создают сооружения из отдельных посторонних частиц, скрепляя их в единое целое. Речь идет о постройке уже упомянутых «домиков» - трубок из отдельных частичек, которые собираются червями на морском дне и укрепляются с помощью специальных «рабочих» органов – преобразованных передних параподий.
Сам процесс сооружения трубки является сложной деятельностью, состоящей из нескольких фаз, адекватно видоизменяющихся в зависимости от таких внешних факторов, как характер грунта и течения, рельеф дна, количество и состав опускающихся на дно частиц, скорость их оседания и т.д. Строительным материалом служат разного рода мелкий обломочный материал (например, кусочки раковин), песчинки, частицы растений и т.п., которые скрепляются специальными выделениями особого «цементирующего органа». Пригодные для употребления частицы выбираются животным, причем в зависимости от его возраста; молодые особи используют лишь мелкие гранулы, более старые – более крупные. Все частицы собираются и прикрепляются друг к другу уже упомянутыми специализированными щупальцами. Полихета Aulophorus carteri, например, строит трубки из спор водных растений, которые она собирает и прикрепляет друг к другу вокруг себя подобно тому, как строится труба из кирпичей (рис.33).
Рис. 33. Постройка многощетинковым червем трубки из спор водных растений (по Грассэ)
Не меньший интерес представляют впервые намечающиеся у многощетинковых червей проявления брачного поведения и агрессивности, а вместе с ними и элементов общения. Конечно, подлинные агрессивность и брачное поведение характеризуются ритуализацией, которая появляется только у головоногих моллюсков и членистоногих, т.е. на низшем уровне перцептивной психики. Все же у представителей рода Nereis удалось наблюдать борьбу между двумя червями. У Nereis pelagica такая борьба может начаться (но не обязательно), если одна особь пытается проникнуть в «домик» другой особи. Иногда вторгшаяся особь кусает владельца «домика» в задний конец тела, и тогда «хозяин» покидает трубку или же, перевернувшись, вступает в борьбу с пришельцем. В других случаях, когда животные встречаются в трубке на полпути, головой к голове, борьба вспыхивает немедленно, и черви борются, расположившись брюшными сторонами и головами друг к другу. Однако при всех обстоятельствах животные не наносят друг другу повреждения. В более ожесточенной, «непримиримой» форме борьба происходит у Nereis caudata, причем не только из-за обладания «домиком». «Драки» описаны английским исследователем поведения морских кольчатых червей С.М.Эвансом и его сотрудниками также у полихеты Harmothoe imbricata. У этого вида борьба может возникнуть при случайной встрече двух особей, особенно же агрессивность проявляется в сфере воспроизведения, при образовании пар: самец, уцепившийся за самку, становится чрезвычайно агрессивным по отношению к другим самцам (но не самкам). Никогда, однако, борьба не сопровождается у полихет подачей каких-либо сигналов или другими проявлениями ритуализации поведения.
Виноградные улитки же выполняют сложные «брачные игры», длящиеся иногда по нескольку часов, во время которых партнеры принимают по отношению друг к другу различные позы, колют друг друга известковыми спикулами («любовными стрелами») и т.д. Только после такой взаимной стимуляции начинается собственно спаривание (перенос сперматофора). У некоторых полихет, например у Platynereis dumerilii, в последние годы также были описаны брачные «танцы», однако, очевидно, еще рано делать выводы о их конкретном значении и о том, имеют ли они какое-то отношение к ритуализованному поведению. Но во всех этих случаях, несомненно, обнаруживаются некие зачатки, предваряющие значительно более сложные формы поведения высших позвоночных.
Пластичность поведения
Поведение кольчатых червей, как и других низших беспозвоночных, характеризуется малой пластичностью, консервативностью. Господствуют врожденные стереотипы («врожденные программы поведения»). Индивидуальный опыт, научение, играет в жизни этих животных еще небольшую роль. Ассоциативные связи формируются у них с трудом и лишь в ограниченных пределах. Результаты научения сохраняются недолго. У всех кольчатых червей встречается наиболее простая форма научения – привыкание, с которым мы уже встречались у простейших, у которых она является главной, если не единственной формой модификации врожденного, видотипичного поведения. У низших многоклеточных животных врожденные реакции на определенные раздражения также прекращаются после многократного повторения, если не последует адекватного подкрепления этих реакций. Так, например, дождевые черви перестают реагировать на повторное затенение, если оно остается без последствий.
Отмечается привыкание и в сфере пищевого поведения: если многощетинкового червя повторно «кормить» комочками бумаги, смоченными соком его обычной жертвы, он перестает их принимать. Если попеременно с такими комочками давать ему подлинные кусочки пищи, то он в конце концов научается различать их и будет отвергать лишь несъедобную бумагу. Подобные опыты ставились и на кишечнополостных (на полипах, находящихся, как мы уже знаем, на более низком уровне элементарной сенсорной психики). При этом полипы вели себя так же, как морские черви: уже после нескольких (до 5) опытов они отбрасывали несъедобные объекты еще до того, как подносили их к ротовому отверстию. Интересно, что нетренированные щупальца этого не делали, даже находясь рядом с тренированными.
Эти эксперименты интересны тем, что показывают способность низших беспозвоночных отличить по побочным физическим качествам съедобное от несъедобного (по вкусу предлагавшиеся объекты были одинаковыми), что подтверждает наличие истинного психического отражения уже на этом, низшем, уровне филогенетического развития. Ведь здесь производится опосредствованное действие, отличающееся тем, что свойство (или сочетание нескольких свойств), которым животное руководствуется при оценке пригодности объекта к пищевому употреблению, выступает как подлинный сигнал, а чувствительность червя или полипа играет роль посредника между организмом и компонентом среды, от которого непосредственно зависит существование животного.
Более сложное научение путем «проб и ошибок» и формирования новой индивидуальной двигательной реакции можно в элементарной форме обнаружить, начиная уже с плоских червей. Так, планарии, встречая на своем пути полоску наждачной бумаги, сперва останавливаются, но затем все же переползают через нее. Если же сочетать соприкосновение червя с наждачной бумагой с его сотрясением, то планарии перестают переползать через нее и тогда, когда не производится сотрясения. Правда, здесь, очевидно, еще нет подлинной ассоциации между двумя раздражителями (шероховатость и сотрясение); скорее всего, происходит суммация первоначально не очень сильного отрицательного раздражения (шероховатость) с дополнительным отрицательным раздражением (сотрясение) и что, вероятно, влечет за собой общее повышение возбудимости животного.
Все же можно выработать уже у планарии и более сложные реакции, когда один из раздражителей является биологически «нейтральным». В таких случаях можно говорить, очевидно, уже об элементарных процессах подлинного ассоциативного научения. Так, например, Л.Г.Воронин и Н.А.Тушмалова сумели выработать у планарии и кольчатых червей оборонительные и пищевые условные рефлексы. При этом были получены и данные, свидетельствующие об усложнении форм временных связей. Если по поводу плоских червей можно говорить лишь о примитивных неустойчивых условных рефлексах, то у многощетинковых червей обнаруживаются уже стабильные самовосстанавливающиеся после угасания условные рефлексы. Эти различия соответствуют глубоким морфологическим различиям в строении нервной системы этих групп животных и свидетельствуют о существенном прогрессе психической активности у полихет.
Пластичность поведения дождевых червей очень убедительно была показана еще в 1912 г. известным американским зоопсихологом Р.Иерксом. В проведенных им опытах черви должны были в Т-образном лабиринте выбирать определенную сторону, где находилось «гнездо» (в противоположной стороне червь получал электрический удар). Чтобы этому научить червей, потребовалось 120– 180 опытов. (Улитки осваивают такую задачу после 60 опытов и запоминают верное решение в течение 30 дней.) Впоследствии аналогичные опыты с дождевыми червями ставились Л.Геком, И.С.Робинсоном, Р.И.Войтусяком (рис. 34) и другими исследователями. Удавалось также переучивать червей, поменяв «гнездо» и электроды местами. Характерно, что результат научения не меняется и после удаления передних сегментов. Более того, даже лишенный с самого начала этих сегментов, червь оказывается способным научиться правильно ориентироваться в лабиринте.
Подобные факты побудили в свое время В.А. Вагнера говорить о «сегментарной психологии» кольчатых червей и других «членистых», имея при этом в виду, что ганглии каждого сегмента в большой степени обеспечивают автономное выполнение элементарных психических функций. Обезглавленные кольчецы, писал Вагнер, «не теряют способности к спонтанным движениям, за исключением тех лишь, которые стоят в прямой зависимости и связи с органами чувств головы... Обезглавленные черви удерживают свои инстинктивные действия, даже те, в которых голова принимает прямое участие».*
* Вагнер В.А. Сегментарная психология. Новые идеи в биологии. Сборник № 6. Биопсихология. Спб., 1914. С. 123.
|
|
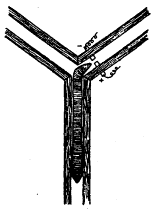
|
|
Рис. 34. Научение у дождевого червя (опыты Войтусяка)
Все это свидетельствует о том, что на уровне даже высших червей цефализация еще не достигла такого развития, чтобы оказать решающее влияние на все поведенческие акты животного. Все же общая направляющая роль головного мозга достаточно велика уже и на этом уровне. Это проявляется, например, в том, что если разрезать пополам дождевого червя, передняя половина будет закапываться в почву, производя вполне координированные движения, задняя же будет лишь беспорядочно извиваться.
Еще лучше, чем у малощетинковых червей, как уже отмечалось, вырабатываются ассоциативные связи у полихет. В опытах удавалось, например, переделать отрицательную реакцию на освещение в положительную путем сочетания освещения с подкармливанием. В итоге черви стали при освещении выползать из своих домиков и без пищевого подкрепления. Время реакции (между световым раздражением и ответным движением) уменьшилось при этом с 28,9 до 3,5 секунды (опыты М.Купленда).
Обстоятельно изучал способность к научению полихет С.М.Эванс. Применяя наказания электрическим током, этот исследователь довольно легко выработал у подопытных животных отрицательную двигательную реакцию (отказ от вползания в узкий темный коридор). В других опытах червям приходилось проползать Т-образный лабиринт. При правильном повороте они могли в течение 5 минут пребывать в затемненной камере. Особенно хорошо обучались полихеты Nereis virens, которым понадобилось менее 100 сочетаний для почти безошибочного решения задачи и ее прочного запоминания; спустя 48 часов они безошибочно воспроизводили правильный путь через лабиринт (рис. 35).
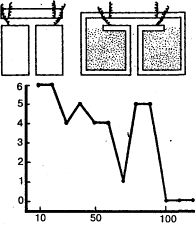
Рис. 35. Научение у Nereis virens (опытЭванса). Вверху даны схемы Т-образных лабиринтов, в которых производились опыты. Внизу – кривая научения (по оси ординат–число ошибок, по оси абсцисс – число опытов)
Общая характеристика высшего уровня элементарной сенсорной психики
Как уже отмечалось, наиболее низкоорганизованные формы многоклеточных беспозвоночных стоят на том же уровне психического развития, что и высшие представители простейших. Но то, что здесь не обнаруживаются существенные различия в поведении, несмотря на глубокие различия в строении, не должно нас удивлять, ибо, как уже говорилось, простейшие олицетворяют собой совершенно особую, рано отклонившуюся филогенетическую ветвь, которая до известных пределов развивалась параллельно ветви низших многоклеточных животных.
Что же касается поведения кольчатых червей, то оно вполне отвечает стадии элементарной сенсорной психики, ибо слагается из движений, ориентированных лишь по отдельным свойствам предметов и явлений (или их сочетаниям), причем это те свойства, которые, по Леонтьеву, оповещают о появлении жизненно важных условий среды, от которых зависит осуществление основных биологических функций животных. Эта ориентация осуществляется, таким образом, на основе одних лишь ощущений. Перцепция, способность к предметному восприятию, еще отсутствует. Не исключено, правда, что у некоторых наземных улиток, как и у упомянутых выше свободно плавающих хищных моллюсков и полихет, уже намечаются зачатки этой способности. Так, виноградная улитка обходит преграду еще до прикосновения к ней, ползет вдоль нее, но только если преграда не слишком велика; если же изображение преграды занимает всю сетчатку, улитка наталкивается на нее. Не реагирует она и на слишком мелкие предметы.
В поведении кольчатых червей еще преобладает избегание неблагоприятных внешних условий, как это имеет место у простейших. Но все же активный поиск положительных раздражителей занимает в поведении кольчецов уже заметное место, и это весьма характерно для высшего уровня элементарной сенсорной психики. Так же как и у простейших, в жизни кольчатых червей и других низших многоклеточных беспозвоночных большую роль играют кинезы и элементарные таксисы. Но наряду с ними здесь уже встречаются зачатки сложных форм инстинктивного поведения (особенно у некоторых многощетинковых червей, пиявок, а также улиток) и впервые появляются высшие таксисы, обеспечивающие значительно более точную и экономную ориентацию животного в пространстве, а тем самым и более полноценное использование пищевых ресурсов в окружающей среде. В результате возникли предпосылки для поднятия всей жизнедеятельности на более высокую ступень, что и имело место на стадии перцептивной психики.
Как было показано, у высших представителей рассматриваемой группы беспозвоночных впервые появляются зачатки конструктивной деятельности, агрессивного поведения, общения. Здесь опять подтверждается правило, что высшие формы поведения зарождаются уже на низших стадиях развития психической деятельности.
Давая общую оценку поведению низших многоклеточных беспозвоночных, следует отметить, что, очевидно, первично главная функция еще примитивной нервной системы состояла в координации внутренних процессов жизнедеятельности в связи со все большей специализацией клеток и новых образований – тканей, из которых строятся все органы и системы многоклеточного организма. «Внешние» же функции нервной системы определяются степенью внешней активности, которая у этих животных находится на еще невысоком уровне, зачастую не более высоком, чем у высших представителей простейших. Вместе с тем строение и функции рецепторов, как и «внешняя» деятельность нервной системы, значительно усложняются у животных, ведущих более активный образ жизни. Особенно это относится к свободноживущим, активно передвигающимся формам.
Говоря об обширной и пестрой по составу группе низших беспозвоночных, нужно, конечно, учесть, что поведение этих животных еще очень слабо изучено. Мы, например, еще почти ничего не знаем об онтогенезе поведения этих животных, о том, как формируется и развивается их поведение, да и совершенствуется ли оно вообще в процессе индивидуального развития, на что уже указывалось в части, посвященной онтогенезу поведения. Вполне возможно (если, конечно, исключить метаморфозные преобразования, личиночное поведение у низших многоклеточных и т.п.), что подобное онтогенетическое совершенствование у обсуждаемых животных не является существенным или даже вообще не происходит, ибо жесткость врожденных программ поведения, исключительная стереотипия форм реагирования являются у этих животных определяющей чертой всего их поведения.
Глава 2 ПЕРЦЕПТИВНАЯ ПСИХИКА
Низший уровень развития перцептивной психики
Перцептивная психика является высшей стадией развития психического отражения. Напомним, что эта стадия характеризуется, по Леонтьеву, изменением строения деятельности – выделением содержания деятельности, относящейся к условиям, в которых дан объект деятельности в среде (операции). По этой причине мы встречаемся здесь уже с подлинными навыками и восприятиями. Предметные компоненты среды отражаются уже как целостные единицы, в то время как при элементарной сенсорной психике имело место отражение лишь отдельных их свойств или суммы последних. Предметное восприятие обязательно предполагает определенную степень обобщения, появляются чувственные представления.
Разумеется, перцептивная психика, свойственная огромному числу животных, стоящих на разных ступенях эволюционного развития, обнаруживает в конкретных своих проявлениях большие различия. Поэтому необходимо и на этой стадии развития психики выделить отдельные уровни, из которых здесь будет рассмотрен низший.
На этом уровне находятся прежде всего высшие беспозвоночные – головоногие моллюски и членистоногие (по меньшей мере высшие). Характеристику рассматриваемого уровня психического развития мы дадим на примере насекомых – центрального класса типа членистоногих. (Кроме насекомых к членистоногим относятся ракообразные, паукообразные, многоножки и некоторые мелкие группы.) Насекомые – самый многочисленный класс животных как по количеству видов, так и по количеству особей. Насекомые обитают повсюду на суше (во всех климатических зонах) – как на поверхности, так и в почве, во всех пресных водоемах, а также в воздухе, поднимаясь на высоту до двух километров.
Движения
Огромное разнообразие «экологических ниш», занимаемых насекомыми и другими членистоногими, обусловило, естественно, развитие весьма различных форм приспособления в области строения и поведения этих животных. Наиболее существенным и характерным для эволюции членистоногих, особенно насекомых, было завоевание новой среды обитания – суши. Членистоногие – первые наземные животные в истории Земли. Переход на сушу был сопряжен с развитием особых органов передвижения – конечностей в виде сложных рычагов, состоящих из отдельных, соединенных суставами члеников, по которым членистоногие и получили свое название. Такие сложные рычаги (их аналогами являются конечности наземных позвоночных) наилучшим образом служат для передвижения по твердому субстрату путем отталкивания от него. Членистоногие в состоянии передвигаться по суше почти всеми возможными способами (ползание, ходьба, бег как по горизонтальным, так и по вертикальным поверхностям, прыгание, передвижение в толще грунта). В воде членистоногие плавают, ныряют, ползают и даже бегают по ее поверхности.
У насекомых, кроме того, в связи с переходом к жизни в воздушной среде впервые появился и совершенно новый способ передвижения – полет с помощью крыльев. Насекомые способны развить в полете очень большую скорость: у некоторых стрекоз она достигает 90 километров в час, у самца овода – даже 122 километра в час. Поистине поразительна частота взмахов крыльев у многих насекомых. Так, мотылек делает лишь 9 ударов в секунду, пчела же – 190, а комнатная муха – 330, но у некоторых мух и перепончатокрылых скорость вибрации крыльев достигает до тысячи ударов в секунду. Столь быстрые движения и соответственно частоты мышечных сокращений не наблюдаются ни у каких других животных.
Соответственно и крылевая мускулатура насекомых отличается исключительно сложным устройством: она состоит из множества различно располагающихся мышечных пучков, частично действующих как антагонисты. Большую сложность обнаруживают и другие отделы мышечной системы.
Вообще у насекомых (не считая крылевой мускулатуры насекомых) различают следующие отделы мышечной системы: мышцы туловища, заведующие его движениями: мышцы конечностей, с помощью которых движется вся конечность по отношению к туловищу; мышцы отдельных члеников конечностей, мышцы внутренних органов.
Как видно, мускулатура насекомых отличается высокой степенью дифференцированности и специализированности. К тому же подавляющее большинство их мышц – поперечнополосатые. Количество отдельных мускулов исключительно велико – полторы тысячи и более. Внушительна и сила мышц: насекомые способны таскать предметы, иногда в сто раз превышающие их собственный вес. В целом, как отмечал Догель, у членистоногих мышечная система беспозвоночных достигает «апогея своего развития» как в морфологическом, так и в физиологическом отношении.
Центральная нервная система
В соответствии со сложной и высокодифференцированной организацией двигательного аппарата находится и сложное строение центральной нервной системы насекомых, которую, однако, мы можем здесь охарактеризовать лишь в самых общих чертах.
Как и у кольчатых червей, основу всей нервной системы составляют у насекомых метамерно расположенные ганглии, связанные друг с другом межсегментными нервными стволами. Эти стволы сближаются у большинства насекомых настолько, что образуют (хотя бы внешне) одну непарную нервную цепочку. Одновременно имеет место и слияние последовательных метамерных ганглиев.
В связи с отчетливо выраженной цефализацией в головном отделе слившиеся ганглии образовали мощный головной мозг в виде сложно устроенного надглоточного узла, в котором различают три отдела: протоцеребрум, дейтоцеребрум и тритоцеребрум. Передний отдел, протоцеребрум, развит сильнее остальных. Особое внимание заслуживают находящиеся в нем так называемые грибовидные (или стебельчатые) тела – ассоциативные и координирующие центры. В наибольшей степени они развиты у насекомых с особенно сложным поведением – у пчел, прежде всего рабочих особей, и муравьев. С протоцеребрумом связана и пара очень крупных и сложно устроенных зрительных долей, иннервирующих сложные глаза. Дейтоцеребрум (средний отдел головного мозга) иннервирует усики, тритоцеребрум иннервирует область рта и, кроме того, тесно связан с симпатической нервной системой. Вообще для насекомых характерна большая концентрация нервных элементов. Такая концентрация повышает возможности управления отдельными системами органов и поднимает общий морфофизиологический уровень организма как целостной единицы.
Нервная система, и в частности головной мозг, устроена у разных членистоногих, однако, весьма различно. Существенные различия обнаруживаются между низшими и высшими представителями даже в пределах классов, а иногда и отрядов. Так, например, отношение размеров мозга к объему туловища равняется у майского жука 1 : 3290, у муравья 1 : 280, а у пчелы 1 : 174.
На очень высоком уровне развития находится нервная система головоногих моллюсков, в корне отличаясь от таковой других моллюсков. Особенно это относится к их весьма сложно устроенному головному мозгу. Обращает на себя внимание исключительная концентрация мозговых ганглиев: ганглии слились воедино и образовали общую околопищеводную массу. Основания зрительных нервов образуют громадные вздутия – оптические ганглии, которые своими размерами нередко превосходят сами церебральные узлы.
Нервы, отходящие от мозга к эффекторам, достигают очень большой мощности – до миллиметра и больше в диаметре, что в 5–10 раз превышает толщину нервов большинства позвоночных! Этим обеспечивается значительно большая скорость проведения нервных импульсов, чем у низших моллюсков. Соответственно огромной величине глаза исключительно развит и зрительный нерв. Выдающийся советский гистолог А.А.Заварзин показал большое сходство тонкого гистологического строения оптических центров головоногих и позвоночных.
Таким образом, мы видим, что глубокие различия между элементарной сенсорной и перцептивной психикой находят свое воплощение и в структуре нервной системы уже на низшем уровне перцептивной психики.
Сенсорные способности, таксисы
Специфические условия, в которых живут насекомые, далеко идущее приспособление двигательной активности при обилии и разнообразии качественно весьма различных агентов среды, управляющих их поведением, – все это обусловило появление у данных животных многих, и притом своеобразно устроенных, органов чувств.
Большую роль в жизни насекомых играет зрение. В разном количестве у разных видов имеются простые одиночные глаза. Но главный орган зрения – своеобразный сложный глаз, получивший название фасеточного, так как его поверхность состоит из отдельных, четко ограниченных участков – фасеток. Каждой фасетке соответствует самостоятельный глазок – омматидий, изолированный от своих соседей пигментной прокладкой и функционирующий независимо от них. Общий внешний покров глаза образует над омматидиями утолщения, обозначаемые как роговица или роговой хрусталик и играющие роль диоптрического аппарата глаза. Каждый омматидий содержит 7– 8 зрительных клеток, аксоны которых оканчиваются в оптических ганглиях мозга.
Благодаря такому устройству сложных глаз в мозг насекомого передаются «мозаичные» изображения воспринимаемых предметных компонентов среды. Конечно, это не означает, что насекомое действительно видит окружающий мир в виде мозаики. В мозгу поступающие из отдельных омматидиев зрительные раздражения синтезируются в целостную картину, как это имеет место и у позвоночных. Все же нам очень трудно представить себе, как на самом деле видит насекомое.
Еще в прошлом веке делались попытки установить путем фотографирования через фасеточные глаза четкость и дальность зрения насекомых и раков. Позже, в частности в 60-х годах нашего века, такие опыты повторялись, но едва ли возможно таким образом решить проблему, ибо, как известно, видит не глаз, а мозг. Насекомые, очевидно, весьма близоруки: ясное видение, например, бабочки-крапивницы возможно лишь на расстоянии приблизительно 2–3 сантиметров. От объектов, расположенных дальше (уже начиная с 5–7 сантиметров), насекомые получают лишь расплывчатые изображения, но, очевидно, этого достаточно для их общей ориентировки в пространстве.
На расстоянии пчела способна увидеть лишь объекты в сто раз большей величины, чем это необходимо для зрения человека. Отдельный цветок она увидит лишь тогда, когда она приблизится к нему вплотную. Все эти особенности зрения насекомых вполне соответствуют тому микроландшафту, в котором живут эти животные.
Острота зрения (разрешающая способность, т.е. способность видеть две сближенные точки) зависит у насекомых при прочих равных условиях, в частности, от количества омматидиев, которое, однако, сильно колеблется даже в пределах отрядов. Так, в отряде жуков это количество колеблется от трех-четырех десятков до нескольких десятков тысяч. У пчелы каждый глаз состоит приблизительно из четырех тысяч фасеток. Чем мельче фасетки, тем слабее в них освещение, но тем детальнее зрение. Особенно большое значение это имеет для предметного зрения. Кроме того, при более выпуклом глазе оси омматидиев сильнее расходятся, и в результате расширяется поле зрения. Одновременно суживается точка, видимая одной фасеткой, чем также увеличивается острота зрения.
Наилучшим образом насекомые видят подвижные объекты или неподвижные предметы во время собственного движения. Пролетающая над лугом пчела, например, видит покачивающиеся на ветру цветы или же изображение неподвижного цветка скользит по ее фасеточному глазу. В любом случае это подвижное (расплывчатое) изображение заставляет ее спуститься вниз и обследовать растение с минимального, т.е. оптимального для зрения пчелы, расстояния.
Что касается цветового зрения, то видимый ими спектр шире, чем у человека, так как его граница проходит в ультрафиолетовом секторе. Спорный вопрос о способности насекомых к восприятию красного цвета в настоящее время, очевидно, решен в положительном смысле (за исключением медоносной пчелы и некоторых других насекомых). Советский энтомолог Г.А. Мазохин-Поршняков считает, что вообще цветовое зрение у взрослых насекомых представляет универсальное явление. По его данным, цвета различаются даже видами, активная жизнь которых протекает ночью.
До сих пор мы касались только реакции насекомых на свет, который играет в их жизни очень большую роль. Не меньшее, если не большее значение имеют для них запахи. В естественном поведении насекомых стимулы различной модальности всегда выступают в комплексе. Так, уже упомянутые осы рода Philanthus охотятся за пчелами, руководствуясь зрением с расстояния 30 сантиметров, но опознают их по запаху лишь с расстояния 15–5 сантиметров. Во время зрительного поиска оса совершенно не реагирует на запах пчелы.
Аналогично обстоит дело и у других членистоногих. Водяные пауки рода Pirata ориентируются прежде всего по вибрации поверхности воды, в непосредственной же близости от жертвы – зрением (рис. 36). Другие пауки узнают о попадании в их сеть насекомого, воспринимая вибрации паутины. Но обследование жертвы производится сразу тремя способами: паук прикасается к ней передними конечностями, снабженными хеморецепторами, а также так называемыми щупиками (органы осязания), и, кроме того, паук на вкус пробует жертву укусом.
Рис. 36. Водяной наук Pirata бежит по поверхности воды к источнику вибрации (центральная точка). Только непосредственно около неподвижной мухи вид последней приобретает преобладающее влияние и паук сворачивает к ней (опыт Берестынской-Вильчек)
Для ознакомления с таксисами у насекомых рассмотрим в качестве примера фототаксисы. Эти таксисы (положительные или отрицательные) встречаются у насекомых прежде всего в виде тропо-, тело- и менотаксисов, хотя не утратили своего значения и примитивные таксисы. Так, например, положительные и отрицательные клинотаксисы с четко выраженными маятникообразными поисковыми движениями играют ведущую роль в оптической ориентации у многих личинок мух. Тропо- и телотаксисы связаны у них друг с другом многими переходными формами. У взрослых насекомых чаще всего встречаются, однако, четко выраженные телотаксисы. Как правило, тропотаксисное движение очень скоро переходит у них в телотаксисное. У комаров и вшей преобладает тропотаксисная ориентация движения.
Менотаксисы, относящиеся к более сложным формам ориентации, встречаются у высших насекомых (муравьев, пчел, бабочек, жуков). Немецкий исследователь поведения насекомых М.Линдауер выделяет здесь следующие четыре случая: 1) угол фиксации источника света случаен (муравьи, жук-навозник); 2) угол фиксации является врожденным (некоторые бабочки); 3) угол фиксации сообщается особями одного вида друг другу («танцы» пчел, информирующие об угле ориентации по положению солнца); 4) угол фиксации выучивается в индивидуальном порядке. Последний случай можно уже отнести к высшей категории таксисов – мнемотаксисам (выученная ориентация), которые широко распространены среди позвоночных, но также встречаются у высших насекомых (пчел, ос, муравьев) (рис. 37).
Рис. 37. Мнемотаксис у роющей осы Philanthus. После отлета осы кольцо из шишек вокруг норки было перемещено в сторому. После возвращения оса ищет норку внутри кольца и находит ее лишь после повторных облетов (опыт Тинбергена)
Необходимо, однако, подчеркнуть, что и в тех случаях, когда ориентация осуществляется на основе других фототаксисов, поведение насекомых одновременно определяется рядом дополнительных факторов. Например, доказано, что у многих насекомых характер их фототаксисного поведения сильно видоизменяется в зависимости от температуры воздуха, времени дня, физиологического состояния и т.д. Хорошо известно, что муха не реагирует на свет, пока спокойно летает по комнате. Но стоит начать ее преследовать, как у нее сейчас же «просыпается» положительный фототелотаксис: она быстро направляется к источнику света, к окну.
Аналогичным образом насекомые реагируют на разнообразные механические, температурные, химические и оптические раздражения, на силу тяжести, и соответственно у них встречаются во многих различных формах тигмо-, рео-, анемо-, гидро-, гео-, термо-, хемо-, фото- и другие таксисы.
Предметное восприятие
Особый интерес представляет способность насекомых (и других членистоногих) к оптическому восприятию форм как необходимого компонента перцептивной психики. На уровне элементарной сенсорной психики различение форм еще невозможно.
До недавнего времени считалось, что насекомые способны к восприятию формы, но лишь в специфических рамках. Эта специфика усматривалась в том, что в экспериментах насекомые, особенно пчелы, оказывались способными различать лишь фигуры, напоминающие по своим очертаниям цветы (например, кресты или звездчатые фигуры), простые же геометрические фигуры они в этих опытах не различали. Однако Мазохин-Поршняков (в работах, проведенных совместно с Г. М. Вишневской) показал, что пчел можно с полным успехом дрессировать и на такие фигуры, как треугольник, круг, квадрат, из чего он заключил, что пчелы способны распознавать фигуры непосредственно по их графическим признакам.
О способности насекомых к распознаванию форм свидетельствуют и полевые опыты Тинбергена, в которых самки роющих ос обучались распознавать круг из сосновых шишек, выложенных вокруг входа в норку. После того как оса улетала за добычей, круг передвигался на 30 сантиметров в сторону. После возвращения оса искала норку сперва в центре круга (рис. 37). В следующих опытах к тому же шишки заменялись черными камешками, а вокруг норки выстраивался треугольник или даже эллипс из шишек, но оса тем не менее прилетала в круг, хотя из предыдущих опытов было известно, что оса вполне способна отличать камешки от шишек. Таким образом, пространственная ориентация осуществлялась здесь только по форме (кругу).
Хотя у высших насекомых способность к предметному восприятию и не вызывает сомнения, в этом отношении они явно уступают позвоночным; Очевидно, это связано со специфическим строением органов зрения: разрешающая способность глаза зависит у насекомых от количества омматидиев. Чем их больше, тем выше эта способность, так как картина воспринимаемого объекта расчленяется на большее число точек. К тому же насекомые, как и другие членистоногие, очевидно, ориентируются преимущественно все же не по предметным компонентам среды как таковым, а по отдельным их признакам, как это имеет место на стадии элементарной сенсорной психики.
У других беспозвоночных, находящихся также на низшем уровне стадии перцептивной психики, – у головоногих моллюсков – зрение играет не меньшую, если не большую роль, чем у насекомых, и является у них, безусловно, ведущей рецепцией (хотя эти животные ориентируются и хорошо развитыми осязанием и химической чувствительностью). Соответственно и орган зрения получил у головоногих исключительное развитие: головоногие обладают наиболее крупными глазами не только среди обитателей морей, но и вообще в животном мире, причем не только в относительном, но и в абсолютном отношении. Глаза гигантских спрутов (род Architeuthus) достигают в поперечнике 40 сантиметров при общей длине животного (считая и «руки») до 20 метров. Для сравнения можно указать, что у всех, даже самых крупных, китов (например, у 30-метрового голубого кита) диаметр глаза составляет немногим больше десяти сантиметров. Поразительна и огромная разрешающая способность (зоркость) глаза головоногих: на один квадратный миллиметр приходится у разных представителей головоногих от 40 000 до 162 000 палочек, у человека – 120 000 – 400 000, у совы, обладающей самым зорким глазом в мире, – 680 000. Что же касается предметного восприятия, то головоногие моллюски в отличие от низших моллюсков уже способны к подлинному предметному восприятию, что выражается прежде всего в различении ими формы объектов. Так, осьминог в состоянии отличить треугольник от квадрата, малый квадрат от большого, крест от квадрата, вертикальный прямоугольник от горизонтального, треугольник от ромба и т.д. Всего, как удалось установить, осьминог способен различать 46 разных форм (опыты Б. Б. Бойкотта и Дж. З. Янга). Все же зрение головоногих, хотя и сближается с таковым позвоночных, отличается и рядом примитивных признаков, делающих отражение предметов действительности недостаточно полноценным. Еще предстоит большая исследовательская работа для выяснения специфических особенностей оптического восприятия головоногих.
Общение и групповое поведение
У высших беспозвоночных уже в полном объеме представлена категория поведения, которая, как было показано, только в зачаточной форме обнаруживается у некоторых представителей низших многоклеточных беспозвоночных. Речь идет об общении, которое у высших беспозвоночных проявляется иногда и в очень сложных формах подлинного группового поведения (в отличие от колониальной жизни, свойственной многим низшим беспозвоночным). Напомним, что общение, в точном значении этого термина, появляется только тогда, когда контакты между животными начинают осуществляться посредством специальных сигнальных действий.
У головоногих моллюсков и членистоногих, особенно у насекомых, мы находим четкие системы коммуникации с передачей информации по разным сенсорным каналам. Это относится прежде всего к пчелам и муравьям, что, конечно, связано со сложной, высокодифференцированной структурой их сообществ и разделением функций между членами последних (распределение пищедобывательнои, защитной и воспроизводительной функций между отдельными особями).
У муравьев разные формы химической чувствительности играют, очевидно, большую роль, чем другие виды рецепции, и общение между особями осуществляется прежде всего по этому каналу: личинки муравьев выделяют специфические вещества, побуждающие взрослых рабочих особей кормить их, по запаху обитатели одного муравейника узнают «своих» и «чужих». Даже о том, жив или мертв муравей, его сородичи узнают по специфическим выделениям.
Согласование поведения обитателей муравейника происходит путем подачи и приема химических сигналов. Достаточно указать на химические средства муравьев (а также пчел и ос), выделение которых служит сигналом тревоги. Эти вещества выделяются муравьями в случае опасности и быстро распространяются по воздуху. Их восприятие вызывает у других муравьев («солдат») защитную реакцию (готовность к нападению на врага), хотя источник опасности им еще не известен. Поскольку, однако, радиус действия такого сигнала, подаваемого одним муравьем (например, в случае вторжения в муравейник какого-либо насекомого), невелик, то он способен указать и место, откуда угрожает опасность. Чем больше эта опасность, тем больше муравьев выделяют химические сигналы тревоги и тем больше, следовательно, особей призывается на помощь.
По другим выделениям, оставляемым на субстрате в виде следа, муравьи узнают, куда, в какую сторону ушел их сородич. К тому же интенсивность такого пахучего следа указывает на количество пищи, обнаруженной в том месте, куда ведет след, тем более что, направляясь к муравейнику, муравьи наносили по пути пахучие метки лишь в тех случаях, когда они находили корм. При этом тем больше оставляется меток, чем больше обнаружено корма. Поскольку эти метки довольно быстро испаряются, вновь идущие по этому следу муравьи будут воспринимать лишь свежие метки, а таких будет все меньше по мере приближения кормового объекта. Таким образом, муравьи, в конце концов узнают о том, что пищевые ресурсы в данном месте исчерпаны, и перестанут к нему направляться.
У некоторых пчел также существует химический способ передачи информации о нахождении пищи. Это относится, прежде всего, к пчелам – из подсемейства Meliponinae, у которых нет «языка танца». Эти широко распространенные в тропиках пчелы не имеют жала, но яростно кусаются. У мелких форм (самая маленькая имеет длину всего 1,5 миллиметра) нашедшая пищу сборщица только приводит других пчел в возбужденное состояние, беспокойно бегая по сотам, толкая их и время от времени подбегая к летку. Вылетающие же пчелы ориентируются только по запаху цветов. У других видов возбуждение пчел первоначально достигается подобным же образом, но дополнительно сборщица, принесшая взяток, приводит соты толчками в вибрирующее состояние. После этого она возвращается к обнаруженным цветам, а на обратном пути оставляет на траве, ветках или камнях через каждые два-три метра пахучие метки, по которым затем приводит группами других пчел к источнику пищи. Это достигается тем, что пчела-разведчица летает взад и вперед между двумя соседними метками, пока другие пчелы не присоединяются к ней. Следовательно, здесь мы имеем дело с комплексной передачей информации по оптическому, тактильно-вибрационному и, возможно, акустическому каналам, но при этом ведущую роль играют оставляемые сборщицей запахи.
У нашей медоносной пчелы также существует химический способ передачи информации, но только как вспомогательный, когда она имеет дело с кормом, лишенным собственного заметного запаха. В этих случаях место нахождения кормового объекта метится секретом из так называемой железы Насонова.
Однако основным и наиболее совершенным способом передачи информации о кормовых объектах являются «танцы». Найдя источник пищи (нектара) и вернувшись в улей, пчела раздает другим пчелам-сборщицам пробы нектара и приступает к «танцу», который состоит из пробежек по сотам. Рисунок танца зависит от местонахождения обнаруженного источника пищи: если он находится рядом с ульем (на расстоянии 2-5 метров от него), то производится «танец-толчок»: пчела беспорядочно бежит по сотам, время от времени виляя брюшком; если же корм обнаружен на расстоянии до 100 метров от улья, то выполняется один из описанных К. Фришем танцев – «круговой», который состоит из пробежек по кругу попеременно по часовой стрелке и против нее (рис. 38). Если же нектар обнаружен на большем расстоянии, то выполняется «виляющий» танец, также описанный и изученный Фришем. Это пробежки по прямой, сопровождающиеся виляющими движениями брюшка с возвращением к исходной точке то слева, то справа (рис. 38, а). Интенсивность виляющих движений указывает на расстояние находки: чем ближе находится кормовой объект, тем интенсивнее выполняется танец (рис. 38, б).
Кроме того, как тоже показал Фриш, во второй форме танца угол между линией пробежки и вертикалью (на вертикально расположенных сотах) соответствует углу между линией полета пчелы от улья к кормовому объекту и положением солнца. Таким образом, горизонтальное направление истинного полета переносится на вертикальную плоскость, ориентация по положению солнца подменяется ориентацией по земному притяжению. При этом, если следует лететь навстречу солнцу, пчела танцует снизу вверх, если же от солнца, то сверху вниз (рис. 38, в). Таким образом, медоносная пчела способна к транспозиции с фотоменотаксисов на геоменотаксисы: фиксированный угол к видимому положению солнца переносится в темном улье к силе тяжести.
Эта способность медоносной пчелы к транспозиции, коренящаяся в широко распространенной среди насекомых способности к переносу менотаксисных движений на вертикальную плоскость (рис. 39), обеспечивает, как мы видим, исключительную полноту и точность передачи информации. Такая транспозиция является частным случаем трансформации, лежащей в основе всякого общения между животными: свойства действительных предметов и ситуации трансформируются на языке животных с помощью видоспецифического «кода» в оптические, акустические и прочие условные сигналы, информирующие других особей об этих свойствах. Несмотря на всю сложность и точность, система коммуникации медоносной пчелы в принципе не отличается от любой другой системы общения у животных. Переводятся ли истинные пространственные параметры в сигналы, воспринимаемые с помощью зрения, осязания и чувства силы тяжести, как это имеет место у медоносной пчелы, или же в сигналы, воспринимаемые хеморецепцией (муравьи), – информация в любом случае передается в трансформированном, условном виде. Здесь как нельзя нагляднее выступают характер и функциональное значение психики у животных.
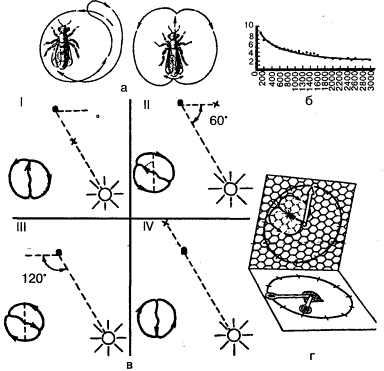
Рис. 38. «Танцы пчел». Обозначения: а – «круговой танец» (слева) и «виляющий» (справа); б – интенсивность выполнения «виляющих танцев» как показатель расстояния до местонахождения источника пищи (на оси ординат отмечено число «танцев» за 15 секунд, на оси абсцисс – расстояние до пищи в метрах); в – «виляющий танец» как показатель направления полета к источнику пищи: I – навстречу солнцу, II – под углом 60˚ налево от положения солнца, III – направо от положения солнца, IV – от солнца. Точка – местонахождение улья, крестик – местонахождение пищи. Изображение «танцев» следует представить себе на вертикальной плоскости (по Фришу); г – схема, иллюстрирующая транспозицию угла полета по отношению к положению солнца (горизонтальная плоскость) на вертикальную плоскость сот, на которых пчела выполняет «виляющий танец» (по Кэстнеру)
Вершиной развития инстинктивных компонентов общения является ритуализация поведения, которая особенно четко выступает в воспроизводительном поведении насекомых, в частности в «ухаживании» самцов за самками. Вот типичный брачный «ритуал» самца бабочки-бархатницы, описанный Тинбергеном: полетев вслед за самкой, самец садится рядом с ней, потом становится перед ней, начинает «дрожать» крыльями (переднезадние движения сомкнутых крыльев), затем повторно раскрывает и закрывает их, что сопровождается постукиванием усиками по крыльям самки. После этого самец обхватывает самку крыльями и зажимает между ними ее усики, которое при этом прикасаются к пахучему участку на крыльях самца. Только после этого, если самка не отвергает «ухаживания» самца, начинается спаривание.
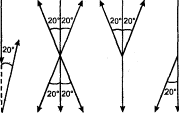
Рис. 39. Транспозиция оптической ориентации в другую сенсорную сферу у разных насекомых. Обозначения:
а – менотаксисное движение по горизонтальной плоскости под углом 20° к источнику света. Вертикальная стрелка – направление светового луча; б–г – продолжение движения по вертикальной плоскости у муравья (б), божьей коровки (в), майского жука (г). Вертикальная стрелка – направление силы тяжести
В этом примере обращает на себя внимание строгая фиксированность последовательности сигнальных поз и телодвижений, что и характерно для ритуализованного поведения. Как мы видим, при ритуализации двигательные акты всегда носят условный характер: выполняемые животным движения несут лишь сигнальную, но не непосредственно «механическую» функцию. Именно на этой основе зародился язык поз, телодвижений и звуков у животных во всем своем удивительном многообразии. При этом в условности сигналов находит непосредственное воплощение психический компонент общения.
Наиболее сложные системы общения присущи, конечно, насекомым, живущим большими семьями. Это относится опять-таки в первую очередь к медоносной пчеле и муравьям. Дифференцированные и многообразные взаимоотношения между членами пчелиной семьи, опирающиеся на сложные формы коммуникации, являются уникальными в животном мире. По поводу этого сообщества высказывались самые различные мнения начиная от антропоморфичного толкования ее как копии человеческого общества или государства (сохранилось неудачное, вводящее в заблуждение название «общественные насекомые») до представления о некоей сумме независимо действующих «рефлекторных автоматов». Вместе с тем оказалось (исследования К. П. Трубецкой), что пчелиная семья представляет собой объединение особей, поведение которых носит самостоятельный, но не независимый характер. Самостоятельность поведения отдельных пчел проявляется прежде всего в резко выраженных индивидуальных различиях: предпочтение отдельных видов ульевых работ, различия в сроках и характере перехода от ульевой деятельности к летной и др. Большую сложность в количественном и качественном аспектах достигают взаимоотношения между обитателями улья, в результате чего функциональная настроенность одних пчел может оказать влияние на других. Рабочие особи с их повышенным уровнем нервной деятельности часто сообщают возбуждение другим, менее активным рабочим пчелам.
Территориальное поведение
Уже у кольчатых червей и низших моллюсков наблюдаются первые проявления территориального поведения, что выражается в приуроченности жизнедеятельности к постоянно занимаемому участку, на котором располагается и убежище («дом») животного. Правда, выполнение жизненных функций (или хотя бы части из них) на определенном участке составляет лишь часть территориального поведения, равно как и наличие «дома» само по себе еще не является критерием последнего. При полноценном территориальном поведении, встречающемся лишь на уровне перцептивной психики, животное-хозяин своим видом, голосом или запахом дает знать сородичам о своем присутствии на занятом участке. Важную роль играет при этом «маркировка местности», т.е. нанесение пахучих или иных меток на разные объекты, особенно на периферии участка.
Такими метками могут быть испражнения или выделения кожных желез, а также оптические знаки – содранная со стволов деревьев кора, вытоптанная трава и т.п.
Весьма существенным признаком территориального поведения является активная защита участка хозяином. Тинберген придает этому критерию наряду с привязанностью к местности особое значение. Враждебное отношение («агрессивность») проявляется хозяином по отношению к любому представителю того же вида, особенно того же пола.
Часто такое поведение приурочено лишь к периоду размножения. Так, например, древесный таракан (Cryptocereus punctulatus), устраивающий гнездовую камеру в гнилой древесине, весьма интенсивно (и обычно успешно) защищает место, где находится эта камера, от вторжения других самцов. В случае поражения гнездовую камеру, где находится и самка, занимает победитель.
В высокоразвитых формах территориальное поведение, как показал немецкий этолог А.Хеймер, встречается у стрекоз. Это интересно уже потому, что стрекозы относятся к самым древним насекомым и, появившись в палеозое, дожили до наших дней, не претерпев за эти 50 миллионов лет существенных морфологических изменений. Вместе с тем, как показывает Хеймер, архаичность строения сочетается у них с высокоразвитым поведением, причем не только в сфере территориальности. Этот, казалось бы, парадоксальный факт сохранения древнейших признаков строения при наличии весьма прогрессивных форм поведения вновь подтверждает общее правило несоответствия морфологических и поведенческих признаков, если иметь в виду морфофункциональные отношения на уровне целого организма.
Что же касается территориального поведения стрекоз (речь идет конкретно о равнокрылых стрекозах-красотках Zygoptera), то половозрелые самцы отыскивают места, пригодные для постоянного пребывания, которые и становятся их индивидуальными участками. Эти участки маркируются оптически, а именно путем ежедневных многократных облетов, и энергично обороняются от самцов-сородичей. Внутри индивидуального участка имеются основные и дополнительные места отдыха, а также зона для откладки яиц, границы которой маркируются упомянутым образом. В эту зону самец приводит самку посредством совместно выполняемого с ней «танца» над поверхностью воды. Кроме того, самец «ухаживает» за самкой, выполняя на своей территории особый ритуализованный брачный полет, а во время откладки яиц он охраняет и сторожит самку или соответственно несколько самок, ибо один самец нередко спаривается подряд с несколькими самками. Охрана проявляется в том, что самец отгоняет других мужских особей, пытающихся спариваться с «его» самками, что мешает откладыванию яиц, сторожевая же функция состоит в том, что при попытке самок прервать это занятие самец заставляет их вернуться и возобновить его. Добавим еще, что территориальная борьба самцов выполняется в высокоритуализованных формах (подлинная борьба отсутствует!). В целом все поведение самцов (как и самок) отличается большой пластичностью.
Таким образом, у данных видов стрекоз существует вполне развитое территориальное поведение: занимаются индивидуальные участки, в которых выделяются функциональные зоны (отдыха, размножения), на этих участках выполняются все жизненные функции (за исключением сна: вечером самцы собираются в особых местах ночевок), хозяин дает знать о своем присутствии, маркирует и активно защищает участок.
Интересные особенности территориального поведения муравьев были выявлены советским энтомологом А. А. Захаровым. Оказалось, что у муравьев существуют два основных типа использования кормовых участков: совместное использование угодий несколькими семьями и использование кормового участка населением лишь одного гнезда. При этом выявилась прямая связь между плотностью муравьев на участке и «агрессивностью»: у видов с низкой плотностью участки не охраняются (кроме пригнездовой зоны), при высокой же плотности на кормовых участках появляются охраняемые территории, а между ними «нейтральные зоны» (рис. 40). Как показал другой советский исследователь, Г. М. Длусский, муравьи того же вида, принадлежащие к другим семьям, равно как и представители других видов, в пределы этих территорий не допускаются.
|
|
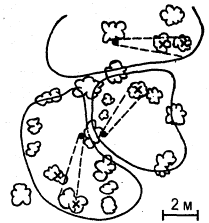
Рис. 40. Территориальное поведение муравьев (Acantholepis melanogaster). Территории трех семей с нейтральными зонами между ними. Точки – гнезда; крестики – колонии тлей, посещаемые муравьями; замкнутые кривые – кустарник; пунктирные линии – основные направления поиска и транспортировки пищи (по Захарову)
Наибольшую сложность территориальное поведение достигло у рыжих лесных муравьев, у которых наблюдается и наибольшая плотность особей на кормовых участках. Охраняемые территории этих муравьев достигают значительно большей величины, чем у других видов. Центральными элементами такой территории являются муравьиные тропы (постоянные кормовые дороги), длина которых нередко превышает 100 метров, а протяженность всей сети троп – 1 километр. Каждой дорогой пользуется лишь определенная группа муравьев («колонна»), занимающая в муравейнике определенный сектор, примыкающий к «своей» дороге. Территория всей семьи подразделяется дорогами на отдельные части, составляющие территории отдельных колонн. Между ними имеются такие же нейтральные пространства, как и между территориями семей, хотя и не столь выраженные, как в последнем случае. Границы территории маркированы и препятствуют «рассеиванию» муравьев. Муравьи двигаются по территории не хаотично, а в соответствии со структурой территории: в центральной и средней ее частях – по кратчайшему пути от дорог или муравейника, на периферии же – параллельно границам территории. Таким образом, траектория движения каждого муравья детерминируется местом нахождения его на территории (исследования И. В. Стебаева). Наибольшую охотничью активность муравьи развивают на периферии территории (далее 10 метров от гнезда). Сбор строительного материала производится в пределах двухметровой полосы вдоль дорог.
Захаров справедливо отмечает сходство территориального поведения муравьев с таковым у птиц и хищных млекопитающих.
Инстинкт и научение в поведении насекомых
Долгие годы господствовало мнение будто насекомые и другие членистоногие являются существами, поведением которых руководит жесткий «слепой инстинкт». Это представление укоренилось преимущественно под влиянием работ выдающегося французского энтомолога Ж. А. Фабра, который сумел своими блестящими исследованиями убедительно показать, что даже сложнейшие действия насекомых не являются проявлением «разума», а выполняются на врожденной, инстинктивной основе. Одностороннее развитие положений Фабра и привело к указанной, неверной оценке поведения насекомых, к отрицанию не только разумности их поведения, но и к отрицанию, или хотя бы умалению роли накопления индивидуального опыта, научения в их жизни.
Как мы уже видели, формирование любой формы видотипичного, наследственно «закодированного», т.е. инстинктивного, поведения в онтогенезе всегда сопряжено в той или иной степени с какими-либо элементами индивидуально приобретаемого поведения, научения. О строго фиксированном инстинктивном поведении в «чистом виде» не приходится говорить даже относительно низших животных.
В полной мере это относится и к насекомым, инстинктивное поведение которых также совершенствуется научением. В этом состоит основная роль научения в жизни насекомых. Можно, очевидно, считать, что научение стоит у насекомых и других членистоногих «на службе» у инстинктивного поведения. Как и у других животных, инстинктивные движения (врожденные двигательные координации) у них генетически строго фиксированы. Инстинктивные же действия, инстинктивное поведение являются и у насекомых в той или иной степени пластичными благодаря включению в них благоприобретаемых компонентов.
В естественных условиях способность к накоплению индивидуального опыта проявляется у насекомых в неодинаковой степени в разных функциональных сферах. Чаще всего она связана с ориентацией в пространстве и пищедобывательной деятельностью. Примером могут служить отмеченные выше опыты по обучению пчел за пищевое подкрепление ориентироваться по различным рисункам. Другой пример – муравьи, которые очень легко (всего за 12–15 опытов) научаются проходить даже сложный лабиринт, но, насколько известно, не научаются действиям, лежащим за пределами указанных функциональных сфер. Такая специфическая направленность (и одновременно ограниченность) способности к научению является характерной особенностью научения у представителей всего типа членистоногих.
Роль научения в поведении насекомых наглядно выступает и в «танцах» пчел – этих высших представителей членистоногих. Отстаивая взгляд, будто насекомые, в том числе пчелы, являются «стимульно связанными, рефлекторными животными», американские ученые В. Детьер и Э. Стеллар заявляют, например, что выполнению и интерпретации сложного танца пчелы не обучаются. Вместе с тем, как показали советские исследователи Н. Г. Лопатина, И. А. Никитина, Е. Г. Чеснокова и другие, процессы научения не только уточняют, но и модифицируют коммуникационные способности пчелы в онтогенезе и расширяют набор сигнальных средств.
Более того, как установили названные исследователи, биологическая значимость сигнальной деятельности медоносных пчел определяется стереотипом условных рефлексов, приобретаемых в онтогенезе по мере освоения пространства и при общении в семье. Оказалось, что интерпретация передаваемой в танце информации о расстоянии и направлении полета к источнику пищи возможна лишь в том случае, если пчела до этого научилась соотносить местонахождение корма с характером информации, содержащейся в танце сборщиц. Кроме того, тактильный компонент танца (вибрации брюшка) не имеет врожденного сигнального значения. Последнее приобретается в онтогенезе также условно-рефлекторным путем: пчелы, не имевшие в онтогенезе контактов (пищевых) с танцовщицей, не в состоянии интерпретировать этот существеннейший элемент танца. -Следовательно, каждая пчела должна в основном научиться «понимать» язык танца. С другой стороны, образование временных связей оказалось важным и для формирования самой способности к выполнению танцев.
Таким образом, нет неизменных форм поведения даже там, где прежде всего требуется стереотипность, – в сигнальных позах и телодвижениях. Даже такое врожденное коммуникативное поведение, как «танцы» пчел, не только дополняется и обогащается процессами научения, не только переплетается с ними, но и формируется в комплексе с индивидуально приобретаемыми элементами поведения.
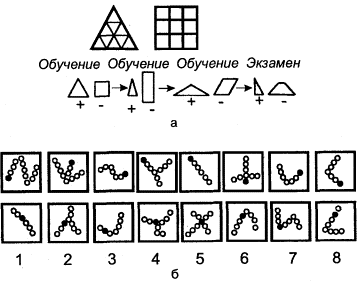
Рис. 41. Изучение способности медоносной пчелы к зрительному обобщению (опыты Мазохина-Поршнякова). Обозначения: а – общая схема проведения опытов; вверху – тестовые фигуры, внизу – последовательность отдельных этапов формирования реакции на обобщенные признаки треугольника и четырехугольника (+ = пищевое подкрепление); б – опознавание рисунков по локальному признаку. В каждом опыте предлагалась на выбор одна пара из верхнего и нижнего ряда рисунков; подкреплялись только фигуры из верхнего ряда
Конечно, медоносная пчела занимает среди насекомых исключительное положение, и далеко не у всех представителей этого огромного класса психическое развитие достигает такой высоты. Об исключительных психических качествах медоносной пчелы свидетельствуют, в частности, экспериментальные данные, говорящие о наличии у нее аналогов некоторых психических функций высших позвоночных. Речь идет об установленной Мазохиным-Поршняковым высокоразвитой способности пчелы к зрительным обобщениям, например, типа «треугольник» и «четырехугольник» (независимо от конкретной формы, соотношения размеров и взаимной ориентации фигур) (рис. 41, а), «двуцветность» и др. В одной из серий опытов пчелам предлагалось выбрать из попарно предъявляемых фигур те, у которых один локальный признак (зачерченный кружочек) находился на конце цепочки из кружочков независимо от длины и формы этих цепей (рис. 41, б). Со всеми предложенными им задачами, даже в наиболее сложных вариантах, пчелы вполне справлялись. При этом отмечалась большая пластичность, нестандартность поведения, что экспериментатор справедливо связывает с непрерывной изменчивостью условий среды (непостоянство освещения, взаимного расположения, формы, окраски и многих других признаков компонентов среды), при которых этим насекомым приходится добывать пищу. Мазохин-Поршняков приходит к выводу, что выбор незнакомого объекта на основе обобщенных зрительных образов (иногда неправильно обозначаемых им как «понятия») есть свидетельство нестандартного использования пчелами индивидуального опыта, его применения в новой ситуации, отличной от обстановки первоначальной выработки соответствующего навыка.
Таким образом, здесь справедливо подчеркивается наличие и значение факта переноса определенного навыка в новую ситуацию и решение сложной задачи на основе индивидуального опыта, зафиксированного в виде обобщенного зрительного представления. В этом отношении мы действительно находим уже у пчел психические способности, аналогичные тем, которые относятся к предпосылкам интеллектуальных действий высших позвоночных животных. Однако одних этих предпосылок недостаточно для интеллектуального поведения, мышления животных, особенно если смотреть на эти высшие психические функции животных как на ступень по направлению к зарождению человеческого сознания. Поэтому описанные способности пчел не могут служить и критерием для признания у них мышления и уж во всяком случае не приходится говорить о наличии у пчелы рассудочной деятельности, хотя бы и в элементарной форме, как толкует результаты своих исследований Мазохин-Поршняков. Признавая у высших животных наличие своеобразных мыслительных способностей, интеллекта, нужно со всей определенностью отдавать себе отчет в том, что рассудок, т.е. разум, сознание как качественно иная категория психического отражения не присущи ни одному из животных, а только человеку.
Общая характеристика низшего уровня перцептивной психики
На низшем уровне перцептивной психики уже представлены все те прогрессивные признаки, которые характеризуют перцептивную психику вообще, но во многих отношениях поведение относящихся сюда животных носит и примитивные черты, сближающие его с поведением нижестоящих животных. Так, основную роль играет ориентация поведения по-прежнему по отдельным свойствам предметов, но не по предметам как таковым: предметное восприятие явно играет еще подчиненную роль в общем поведении. Равным образом в последнем преобладают ригидные, «жестко запрограммированные» элементы поведения над гибкими, благоприобретаемыми и т.д.
С другой стороны, мы констатируем на этом уровне четко выраженный активный поиск положительных раздражителей, т.е. положительное таксисное поведение получает мощное развитие. Налицо все виды высших таксисов, включая мнемотаксисы. Последние играют в пространственной ориентации особенно существенную роль, и именно в индивидуальном заучивании ориентиров проявляется в наибольшей степени способность к благоприобретаемому изменению видового поведения, к научению.
Вместе с тем важно подчеркнуть, что хотя у рассматриваемых здесь животных, в частности насекомых, накопление индивидуального опыта, научение играют существенную роль, наблюдается и определенная противоречивость в процессах научения, сочетания прогрессивных и примитивных черт. Специфическая направленность, приуроченность этих процессов к определенным функциональным сферам, как и само подчиненное положение, которое занимает научение по отношению к инстинктивному поведению, несомненно, указывают на переходное положение данного уровня психического развития между элементарной сенсорной и развитой перцептивной психикой.
Но, как мы видели, это никоим образом не означает, что насекомым, как и другим представителям рассматриваемой группы животных, недостает пластичности поведения. Наоборот, и здесь в полной мере проявляется общая закономерность, что усложнение инстинктивного поведения неизбежно сочетается с усложнением процессов научения (и наоборот). Только такое сочетание обеспечивает подлинный прогресс психической деятельности.
Инстинктивное поведение представлено на рассматриваемом уровне психического развития уже весьма развитыми новыми категориями: групповое поведение, общение, ритуализация. Особую сложность приобретают формы общения у видов, живущих огромными семьями, из которых лучше всего изучены пчелы. Язык пчел, этих высших представителей членистоногих, относится к наиболее сложным формам общения, которые вообще существуют в животном мире. Возможно, что новые исследования познакомят нас с поразительными психическими способностями и других насекомых, но на сегодняшний день пчелы представляются нам наиболее развитыми в этом отношении. Наиболее сложные формы инстинктивного поведения закономерно сочетаются у них с наиболее разнообразными и сложными проявлениями научения, что обеспечивает не только исключительную согласованность действий всех членов пчелиной семьи, но и максимальную пластичность поведения особи. Психические способности пчел (как и некоторых других высших насекомых) в некоторых отношениях, очевидно, уже выходят за рамки низшего уровня перцептивной психики.
В ином направлении, чем у членистоногих, шло развитие психической активности у головоногих моллюсков. По некоторым признакам они приблизились к ветви, ведущей к позвоночным, о чем уже свидетельствуют их крупные размеры и отмеченные выше особенности строения нервной системы и особенно зрительного рецептора, что непосредственно связано с резким увеличением скорости движения по сравнению с другими моллюсками.
Поведение головоногих еще совершенно недостаточно изучено, но уже известны многие примечательные их способности. Прежде всего они выделяются существенным усложнением инстинктивного поведения. У головоногих уже встречаются территориальное поведение (занятие и защита индивидуальных участков), «агрессивность», которая только намечается у высших червей, групповое поведение (стайная жизнь кальмаров и каракатиц), в сфере размножения появляются ритуализованные формы поведения, что находит свое воплощение в видоспецифическом «ухаживании» самцов за самками. Все это присуще только высшим животным, помимо головоногих, членистоногим и позвоночным.
Рядом исследователей обращалось особое внимание на весьма развитое у осьминогов «любопытство», что выражается в обследовании ими биологически «бесполезных» предметов, а также на их высокоразвитые манипуляционные и конструктивные способности. Эти способности проявляются в строительстве валов и убежищ из камней, панцирей крабов, раковин устриц и т.п. Этот строительный материал осьминог подбирает, переносит и укрепляет «руками». Иногда такие гнезда представляют собой закрытые со всех сторон строения. По некоторым, правда еще не подтвержденным, наблюдениям, осьминоги способны и к орудийным действиям, пользуясь камнями для защиты.
Очень важным представляется и то обстоятельство, что впервые у головоногих появляется способность к установлению контактов с человеком, к общению с ним, результатом чего является возможность подлинного приручения этих животных (в отличие от насекомых!).
Таким образом, головоногие, оставив далеко позади других моллюсков, как и вообще всех низших беспозвоночных, достигли, несомненно, высокого уровня психического развития, во многом сблизившись с позвоночными животными.
Вместе с тем и у головоногих наблюдается та же противоречивость в способности к научению, что и у насекомых. Так, например, у осьминога вообще хорошо развита способность к научению на зрительные и тактильные стимулы, но в ряде случаев он оказывается не в состоянии решить, казалось бы, несложные задачи. Особенно это относится к преодолению преград: осьминог неспособен находить обходной путь, если приманка (краб) располагается за прозрачной преградой (в стеклянном цилиндре или за проволочной сеткой); Тщетно пытаясь овладеть приманкой в прямом направлении, осьминог не хватает ее сверху, через край. Правда, по Бойтендайку, некоторые осьминоги все же способны решить несложные задачи обходного пути. При этом, очевидно, большое значение имеет прежний опыт особи. Другие головоногие уступают осьминогу по своим психическим способностям.
Конечно, при оценке подобных экспериментов необходимо иметь в виду, что здесь ставятся биологически неадекватные, а поэтому и неразрешимые задачи: в естественных условиях осьминог никогда не оказывается в ситуации, когда непосредственно зримая жертва оказывается недосягаемой. К тому же задачи обходного пути относятся к разряду весьма сложных – с ними не справляются не только черепахи, но и куры (в биологически несравненно более адекватных условиях). Тем не менее следует думать, что в психической деятельности головоногих действительно сочетаются прогрессивные черты, сближающие их с позвоночными, с примитивными – наследством низших моллюсков. К примитивным чертам относится и известный «негативизм» научения: головоногие легче научаются избегать неприятных раздражений, чем находить благоприятные. И в этом нетрудно усмотреть общность с поведением животных, обладающих элементарной сенсорной психикой.
На низшем уровне перцептивной психики находится, очевидно, хотя бы отчасти и ряд представителей низших позвоночных. Однако в корне различные строение и образ жизни членистоногих и позвоночных являются причиной того, что и их поведение и психика, в сущности, несопоставимы. Так, одной из отличительных особенностей насекомых являются их малые по сравнению с позвоночными размеры. В связи с этим окружающий насекомое мир представляет собой нечто совершенно особое: это не микромир простейших, но и не макромир позвоночных. Человеку трудно себе представить этот мир насекомых с его (с нашей точки зрения) микроландшафтами, микроклиматами и т.д. Хотя насекомые живут рядом и вместе с нами, они живут в совершенно иных условиях температуры, освещения и т.п. Уже поэтому психическое отражение действительности не может у насекомых не быть принципиально иным, чем у позвоночных, да и у большинства других беспозвоночных.
Но поскольку наиболее общие признаки психического отражения, характерные для данного его уровня, присущи всем упомянутым животным, можно, очевидно, по поводу рассмотренных нами в качестве примера насекомых сказать, что мы имеем здесь дело с типичными проявлениями низшего уровня перцептивной психики, но в формах, отвечающих тем особым условиям жизни этих животных, которые отмечались выше.
Высший уровень развития перцептивной психики
В мире животных процесс эволюции привел к трем вершинам: позвоночные, насекомые и головоногие моллюски. Соответственно высокому уровню строения и жизнедеятельности этих животных мы наблюдаем у них и наиболее сложные формы поведения и психического отражения. Представители всех трех «вершин» способны к предметному восприятию, хотя, очевидно, только у позвоночных эта способность получила полное развитие. У остальных двух групп перцепция развивалась своеобразными путями и качественно отличается от таковой у позвоночных. Аналогично обстоит дело и с другими решающими критериями стадии перцептивной психики, не говоря уже о том, что высшего уровня перцептивной психики достигли в процессе эволюции вообще только представители позвоночных, и то явно не все. Только у высших позвоночных обнаруживаются все наиболее сложные проявления психической деятельности, которые вообще встречаются в мире животных.
Сравнивая беспозвоночных с позвоночными, необходимо также учесть, что ни головоногие, ни членистоногие не имеют никакого отношения к предкам позвоночных. Путь, ведущий к этим вершинам, отклонился от пути к третьей вершине еще на очень раннем этапе эволюции животного мира. Поэтому высокое развитие морфологических и поведенческих признаков, отмеченных нами у этих животных, является при сравнении с позвоночными лишь аналогией, обусловленной мощным повышением общего уровня жизнедеятельности, характерным для всех трех групп животных.
С филогенетической точки зрения для нас значительно больший интерес могли бы представить иглокожие, которые так же, как и позвоночные, относятся к вторичноротым в отличие от первичноротых, у которых центральная нервная система располагается на брюшной стороне тела и к которым, в частности, относятся моллюски и членистоногие. Однако и современные иглокожие (морские ежи, звезды и т.д.) являются не предками позвоночных, а лишь боковой ветвью представленной весьма специализированными формами с примитивным поведением, характерным для элементарной сенсорной психики.
На этом уровне находятся и низшие хордовые, которые вместе с позвоночными (или «черепными») составляют тип хордовых. К низшим хордовым относятся оболочники и бесчерепные. Оболочники – морские животные, часть которых ведет неподвижную жизнь (асуи-дия). Бесчерепные представлены всего двумя семействами с тремя родами мелких морских животных, наиболее известное из которых – ланцетник. У бесчерепных и позвоночных общим признаком является внутренний осевой скелет, имеющий вид сплошного (хорда ланцетника) или членистого (позвоночник) стержня, над которым располагается центральная нервная система, имеющая форму трубки. Позвоночные подразделяются на классы круглоротых (миноги и миксины), рыб, земноводных, пресмыкающихся, птиц, млекопитающих. К высшим позвоночным относятся только последние два класса, в пределах которых, следовательно, и обнаруживаются проявления высших психических способностей животных. Что же касается низших позвоночных, то здесь намечаются промежуточные ступени психического развития, характеризующиеся разными сочетаниями элементов низшего и высшего уровней перцептивной психики.
У позвоночных нервная трубка образует в головном отделе вздутия, превращающиеся в ходе эмбриогенеза в головной мозг. Уже у наиболее примитивных позвоночных, у круглоротых, имеются все пять отделов головного мозга (продолговатый, задний, средний, промежуточный и передний мозг). Процесс дифференциации и прогрессивного развития мозговых структур достигает, как известно, своей вершины у млекопитающих, причем не только в переднем мозгу (большие полушария и их кора), но и в стволовой части головного мозга, где формируются, в частности, центры высших форм инстинктивного поведения.
Что касается млекопитающих, то они вообще занимают особое место в эволюции животного мира, ибо, как указывал Северцов, «млекопитающие очень редко приспособляются к быстро наступающим переменам в окружающей среде (например, к новым врагам, к новой добыче и т.д.) обычным путем, то есть путем медленного изменения своих органов и их функций. Гораздо чаще это происходит путем быстрого изменения прежних привычек и навыков и образования новых, приспособленных к новым условиям среды. Здесь впервые выступает на сцену совершенно новый и необычайно важный фактор адаптивной эволюции позвоночных животных, а именно их психика».*
* Северцов А.Н. Главные направления эволюционного процесса. Морфобиологическая теория эволюции. 3-е изд. М., 1967. С. 115.
В первой и второй частях этой книги рассматривалось преимущественно поведение высших позвоночных. По этой причине мы ограничимся здесь лишь некоторыми дополнениями к сказанному.
Локомоция
Мышечная система позвоночных состоит из симметрично расположенных вдоль тела сегментов. Правда, посегментное расположение мышц выражено у хордовых слабее, чем у членистоногих и червей, а у высших представителей позвоночных оно сильно нарушено. Различают соматическую мускулатуру, обслуживающую эффекторные органы, и висцеральную – мускулатуру внутренних органов и кожи. Соматическая мускулатура состоит всегда из поперечно-полосатых мышечных волокон.
Конечности позвоночных представлены непарными и парными образованиями. Непарные конечности имеются только у круглоротых и рыб. Это спинной, заднепроходный и хвостовой плавники. Парные конечности, передние и задние, сильно различаются у разных позвоночных соответственно своей функции (плавники, крылья, лапы, ласты, ноги, руки), однако сравнительное изучение строения их скелета позволяет отчетливо проследить эволюционные преобразования от общей примитивной начальной формы.
Основная функция конечностей состоит у всех животных в локомоции, в перемещении животного в пространстве. Однако у многих членистоногих и позвоночных сюда добавляется еще функция опоры приподнятого над субстратом тела. Поэтому в этих случаях говорят об опорно-локомоторной функции конечностей. Не вдаваясь здесь в детальное рассмотрение разнообразных форм этой основной функции конечностей позвоночных, укажем лишь на некоторые существенные моменты.
Видный советский ученый Н.А.Бернштейн писал, что постепенно назревшая в филогенезе потребность в быстрых и мощных движениях – привела на одной из его ступеней к возникновению и параллельному развитию «костно-суставных кинематических цепей скелета» и поперечнополосатой мускулатуры, снабженной соответственными нервными образованиями. У позвоночных эти скелетно-мышечные системы («неокинетические системы», по Бернштейну) получают существенное прогрессивное развитие по сравнению с членистоногими, причем особенно это относится к рассматриваемым здесь высшим позвоночным, в частности к их локомоторным способностям. Бернштейн указывает в этом отношении на глубокие качественные различия между низшими и высшими позвоночными, обусловленные усложнением возникающих перед организмом двигательных задач, возрастанием разнообразия реакций, требующихся от организма, более высокими требованиями в отношении дифференцированности и точности движений. «Достаточно напомнить, – пишет Бернштейн, – насколько, например, аэродинамический полет птицы сложнее почти полностью гидростатического плавания рыбы или насколько богаче по контингентам участвующих движений охота хищного млекопитающего по сравнению с охотой акулы. Молодая отрасль проворных теплокровных млекопитающих победила тугоподвижных юрских завров именно своей более совершенной моторикой».*
* Берншгейн Н.А. О построении движений. М., 1947. С. 9.
В современных этологических исследованиях локомоторная активность изучается в ее видотипичных проявлениях как адаптация к специфическим условиям существования: разновидности и особенности ходьбы, бега, прыганья, лазанья, плавания, полета и т.д. определяются особенностями образа жизни и служат важным приспособлением к окружающей среде. Вместе с тем всем формам локомоции присуща ритмичность, выражающаяся в том, что движения выполняются в четкой последовательности многократно и относительно стереотипным образом (хотя поведение животного в целом при этом не является стереотипным). Эта ритмичность основана на эндогенной центрально-нервной стимуляции и проприоцепторной обратной связи. Помимо проприоцепторной чувствительности внешние импульсы лишь регулируют эти ритмы, соотносят их параметры (сила, скорость, длительность движений и т.д.) с конкретными условиями тех ситуаций, в которых оказывается животное. В частности, внешние раздражения вызывают начало или конец локомоторных движений, хотя и это может произойти в результате эндогенной стимуляции.
Сказанное достаточно поясняет тот факт, что локомоторные движения относятся к наиболее «автоматизированным» и однообразно выполняемым компонентам всей двигательной сферы животных. В этой же связи стоит и относительная малочисленность форм локомоции у каждого вида. Определяющей для локомоции является ее физическая, механическая функция. Сами локомоторные движения дают животному только минимальную информацию об окружающем мире.
При этом необходимо, правда, учесть, что локомоторная активность включает в себя и ориентировочные компоненты, имеющие, конечно, и определенное познавательнее значение. Так, например, прыгающие животные, особенно древесные, должны перед прыжком точно «рассчитать» расстояние. Как показали советские исследователи поведения животных В. М. Смирин и О. Ю. Орлов, это делается с помощью особых движений «взятия параллакса» (рис. 42). Оказавшись в новом месте, летяга «прицеливается» к разным предметам, это же она делает перед каждым прыжком, хотя со временем число таких движений уменьшается. В итоге уходящее от опасности животное придерживается заранее «отработанного» пути без лишних движений и совершает прыжки с поразительной точностью.
|
|

Рис. 42. Специфические движения летяги для определения удаленности предметов. Летяга «прицеливается» перед прыжком: приподнявшись, животное производит боковые движения головой в горизонтальной плоскости. Подобные ориентирующие движения животные производят особенно часто при передвижении в незнакомой местности (по Смирину и Орлову)
Манипулирование
Эффекторные органы позвоночных всегда выполняют сразу несколько функций – помимо основной функции еще большее или меньшее число дополнительных. В этом проявляется мультифункциональность этих органов. Что касается конечностей высших позвоночных, то, как уже отмечалось, их основой является опорно-локомоторная функция: все же дополнительные их функции сводятся к разнообразным формам манипулирования предметами (пищевыми или несъедобными). Особый интерес представляют для зоопсихолога формы манипулирования передними конечностями, которые в процессе эволюции привели к орудийной деятельности приматов и тем самым стали важнейшей биологической предпосылкой зарождения трудовых действий у древнейших людей.
Говоря о манипуляциях, дополнительных функциях передних конечностей у высших позвоночных, необходимо иметь в виду, что сложные и разнообразные дополнительные функции присущи у этих животных и ротовому аппарату, причем существует далеко идущее функциональное взаимодействие между этими двумя основными эффекторными системами. Поэтому целесообразно анализировать дополнительные функции передних конечностей и ротового аппарата в комплексе. Так, например, у взрослых лисиц обнаруживается 45 дополнительных функций передних конечностей и ротового аппарата, у барсука – 50, у енота – около 80, а у низших обезьян (макаков и павианов) – свыше 150 (данные Фабри).
Важно отметить, что по мере специализации млекопитающих в сфере локомоции часть дополнительных функций передних конечностей передается челюстному аппарату. Так, например, в ряду медведь – енот – барсук – лисица конечности лисицы в наибольшей степени приспособлены к продолжительному быстрому бегу. Но при этом дополнительные (манипуляторные) функции конечностей сводятся к минимуму и соответствующие жизненно необходимые движения (например, в сфере питания) выполняются почти исключительно ротовым аппаратом. Как раз наоборот обстоит дело у медведей.
Если иметь в виду познавательное значение манипулирования, то следует особенно выделить те его формы, при которых животное удерживает предмет перед собой свободно на весу обеими или одной конечностью. Такая фиксация предмета дает животному наилучшие возможности для комплексного (оптического, ольфакторного, тактильного, кинестетического и пр.) обследования, особенно в ходе деструктивного воздействия на него (практического анализа). По данным, полученным Фабри, это дает при известных условиях обезьянам, а отчасти и другим способным на это млекопитающим наиболее всестороннюю и разнообразную информацию, необходимую для развития высших форм психической активности. Оказалось, что медведи владеют тремя способами фиксации объекта на весу, еноты – шестью, низшие обезьяны и полуобезьяны – тремя десятками таких способов! К тому же только обезьяны обладают достаточно различными двигательными возможностями, чтобы произвести подлинный деструктивный анализ (расчленение) объекта на весу.
О формировании манипуляционной активности уже говорилось раньше (см. ч. II). Напомним лишь, что первым начинает функционировать челюстной аппарат, позже появляются движения передних конечностей, которые, однако, вначале еще нуждаются в поддержке более сильной системы челюстного аппарата. Впоследствии движения конечностей становятся все более сильными и самостоятельными и одновременно все более разнообразными, т.е. возрастает их мультифункциональность. Ряд функций, в частности хватательные, первоначально выполнявшиеся челюстями, переходит при этом к конечностям. Вершиной этого развития являются дифференцированные движения, выполняемые только одной конечностью. Как уже отмечалось, прогрессивное развитие дополнительных двигательных функций передних конечностей являлось и является важнейшим фактором психического развития.
Комфортное поведение. Сон и покой
Разновидностью манипулирования можно отчасти считать комфортное поведение, служащее уходу за телом животного, с той лишь особенностью, что объектом манипулирования является не посторонний предмет, а именно собственное тело. Но, кроме того, сюда относятся и нелокализованные движения, производимые всем телом и лишенные специальной пространственной направленности. В итоге можно выделить следующие категории комфортных движений: очищение тела, потряхивание, почесывание (определенного участка тела об субстрат), катание по субстрату, купание (в воде, песке и т.д.).
Комфортное поведение широко распространено и среди членистоногих (достаточно вспомнить, как муха чистит лапками голову, крылья и т.п.), иногда встречаются и специальные морфологические образования или даже органы для очищения тела. Инстинктивная природа комфортных движений обнаруживается в видоспецифичности их выполнения, равно как в строгой фиксированности участков тела, очищаемых тем или иным органом и способом. Столь же четко видотипичными являются и позы сна и покоя. Так, например, у зубров, бизонов и их гибридов было описано всего 107 видотипичных поз и телодвижений по 8 сферам поведения (исследования М. А. Дерягиной). Наибольшее число поз (2/3) приходится на сферы сна, покоя и комфорта. При этом любопытно, что у детенышей (до 2–3 месяцев) всех трех изученных групп животных видотипичные различия в этих сферах еще не выражены. Напрашивается вывод, что эти различия формируются постепенно в более позднем возрасте.
Если рассмотреть одну из форм комфортного поведения, облизывание, то, согласно Дерягиной, оказывается, что только бизон облизывает стоя корень хвоста или приподнятую переднюю ногу, но только зубр облизывает стоя бедра задней ноги и т.д. Если не считать гибридов, то существует несколько способов облизывания, из них присущих только зубру – один, только бизону – четыре, общих зубру и бизону – четыре. Здесь опять проявляются четкие различия в инстинктивных движениях у близкородственных видов (зубр и бизон рассматриваются иногда даже как подвиды).
Сенсорные особенности
В полном соответствии с высокоразвитыми эффекторными системами высших позвоночных находится и высокий уровень их сенсорных способностей. Наибольшее значение имеют органы слуха и равновесия (внутреннее ухо, начиная с земноводных – дополнительно среднее ухо, а у млекопитающих также и наружное ухо), обоняния (обонятельные мешки и раковины) и зрения (глазные яблоки). Хорошо развиты также кожная и мышечная (тактильно-кинестетическая) и термическая чувствительность, вкус, а в ряде случаев и другие виды чувствительности (электрическая, вибрационная и др.). Соответственно развиты у позвоночных и разные таксисы.
Сопоставление роли зрения и обоняния в жизни высших позвоночных показывает, что у большинства млекопитающих ведущую роль играет обоняние, которое достигает у них удивительной остроты (только акулы могут в этом отношении сравниваться с ними). Так, крот чувствует добычу сквозь толщу земли, белый медведь – сквозь лед. Бурый медведь чувствует на расстоянии 20 метров мед, зарытый в землю на глубину 50 сантиметров. Хорошо известны поразительные обонятельные способности собак и копытных.
Однако, как уже отмечалось, в каждом конкретном случае степень развития обоняния (и соответствующих морфологических структур в рецепторе и головном мозге) определяется особенностями биологии вида. Так, например, по Б.С.Матвееву, даже у родственных видов, обитающих в сходных условиях, но различающихся по биологии, – у обыкновенной полевки (Mictorus arvalis) и полевой мыши (Apodemus agrarius) – наблюдаются существенные сенсорные различия. Полевые мыши поедают разнообразный корм – много семян, а также животный корм, полевки же – преимущественно зеленый корм, который всегда имеется в избытке. Вследствие этого обоняние развито у полевой мыши сильнее, чем у полевки.
Зрение лучше всего развито у птиц и приматов. Оно играет важную роль в их пищедобывательной, оборонительной, воспроизводительной и других формах поведения, обеспечивая четкую ориентацию животного в пространстве. Особое значение приобретает зрение при дальней ориентации мигрирующих птиц, а также внутритерриториальной ориентации млекопитающих. Хорошо изучена, например, ориентация птиц по топографическим признакам (например, береговой линии), поляризованному освещению небосвода и астрономическим ориентирам – солнцу, звездам. Последний случай представляет собой наглядный пример ориентации на основе менотаксисов.
У многих млекопитающих ориентация на своей территории (индивидуальном участке или территории стада) осуществляется с помощью фото- и хёмомнемотаксисов (по оптическим и ольфакторным меткам). Кроме того, даже у хуже видящих млекопитающих (например, куньих) зрение играет немаловажную роль при ориентации по подвижным объектам, особенно во время ловли добычи. Даже животные с определенно плохим зрением, как, например, моржи, которые над водой близоруки, пытаются ориентироваться с помощью не только обоняния, но и зрения, особенно опять-таки когда имеют дело с подвижными объектами (приближающимися к ним животными или людьми).
Наиболее мощным зрением, очевидно, во всем мире животных обладают хищные птицы. Достаточно сказать, что сокол способен при благоприятных условиях увидеть сидящего голубя на расстоянии почти полутора километров. Равным образом грифы находят свой корм (трупы животных) на огромном удалении с помощью зрения. Но они не в состоянии обнаружить прикрытые останки животных. Но это относится лишь к птицам, обитающим в степных просторах Восточного полушария. Грифы же Центральной и Южной Америки (например, черные американские грифы), разыскивающие пищу в густых лесах, не могут пользоваться зрением и находят ее, как это ни кажется невероятным, не менее успешно и с не меньшего расстояния с помощью обоняния. Аналогично обстоит дело и у млекопитающих. У копытных, обитающих на открытых пространствах, сильнее развито зрение, чем у их родственников, живущих в лесах, даже если животные принадлежат к одному и тому же роду (например, бизон – преимущественно степное животное и зубр – обитатель лесов).
Все эти примеры наглядно показывают, что образ жизни, биология всецело определяют характер поведения и психики животных (в отличие от человека, который благодаря своей общественно-трудовой деятельности достиг далеко идущей независимости от биологических факторов и психическая деятельность которого обусловливается социальными условиями его жизни). В приведенных примеpax это положение материалистической зоопсихологии относится конкретно к сенсорной сфере животных, в частности к вопросу о ведущей рецепции. Как мы видели, в каждом отдельном случае, даже у близкородственных видов, она определяется конкретными условиями жизни животного. Но то же самое относится и к психическому отражению в целом, характер которого также всегда определяется этими условиями.
Зрительные обобщения и представления
Подлинная рецепция, истинное восприятие предметных компонентов среды как таковых возможны лишь на основе достаточно развитой способности к анализу и обобщению, ибо только это позволяет полноценно узнавать постоянно меняющие свой вид (и другие свойства) предметные компоненты среды.
Исследования, проведенные на разных видах позвоночных (помимо круглоротых), показали, что все они способны к предметному восприятию, в частности к восприятию форм. Уже на этом основании можно заключить, что все позвоночные находятся на стадии перцептивной психики. Однако внутри этой стадии наблюдаются существенные различия между низшими и высшими позвоночными.
Так, рыбы способны в эксперименте отличать геометрические фигуры, например квадрат от треугольника, независимо от меняющейся величины этих фигур. Но достаточно в контрольном опыте перевернуть треугольник (острием вниз) или квадрат (придать ему положение ромба), как рыба перестает узнавать эти фигуры. Следовательно, восприятие и узнавание формы оказываются здесь ограниченными, недостаточно гибкими. Отсутствует способность к быстрому переносу сформировавшегося зрительного представления на основе обобщения. Млекопитающие же вполне способны к такому обобщению и легко узнают треугольник или другую геометрическую фигуру любой величины в любом положении. Существенно превосходят рыб также птицы. Правда, и курица, по некоторым данным, не узнает перевернутый треугольник, который до этого безошибочно отличала от другой геометрической фигуры. Вместе с тем курица способна к значительным обобщениям, опознавая, например, геометрические фигуры независимо от их цвета. Более того, будучи обучена выбирать сплошной треугольник, она также реагирует на него, когда он предъявляется ей в виде штриховки или даже лишь намечен тремя точками! Так же как и рыбы, птицы способны распознавать относительную величину фигур и так же ошибаются на основе зрительных иллюзий.
Способность к зрительному обобщению у многих позвоночных экспериментально доказана. Особенно в отношении внешних их представителей было показано, что они в состоянии даже в очень сложных ситуациях вычленять существенные детали в воспринимаемых объектах и узнавать эти объекты в сильно измененном виде. Отсюда напрашивается вывод о существовании у позвоночных достаточно сложных общих представлений. Представления чрезвычайно важны для выживания. В них закрепляется в общем виде индивидуальный опыт, что позволяет животному легче ориентироваться в существенно меняющейся окружающей среде при временном отсутствии жизненно важных раздражителей. Подобные зрительные обобщения обнаруживаются, в частности, в явлениях «экстраполяции», описанных Л. В. Крушинским, которые проявляются в том, что в ряде случаев животные способны, проследив за движением какого-либо предмета, ориентироваться затем на ожидаемый путь его перемещения. Следовательно, учитывается предстоящее передвижение объекта после его исчезновения. В проведенных им опытах птицы и млекопитающие помещались перед туннелем, имевшим посредине разрыв, через который можно было наблюдать за движением перемещающейся в туннеле кормушки с приманкой. В других опытах применялась ширма со щелью. Некоторые из подопытных животных (врановые, хищные) обегали затем туннель или ширму по направлению движения приманки и ожидали ее в месте появления. На такое «предвосхищающее» поведение оказались, однако, способными лишь животные, имеющие в естественных условиях дело с прячущейся добычей, которую приходится выслеживать или преследовать, зачастую обгоняя ее, или же сами прячущие корм про запас. Как уже раньше указывалось, мы имеем здесь дело с широко распространенной среди высших позвоночных способностью к нахождению обходного пути. При этом важно отметить, что в любом случае, когда животное огибает непрозрачную преграду или даже просто отворачивается от целевого объекта, оно на время теряет последний из виду. Более того, каждое представление по самой природе своей требует отсутствия ранее воспринятых объектов.
Убедительные доказательства тому, что обобщенные зрительные образы в форме представлений широко распространены среди позвоночных, дали эксперименты, проведенные методом отсроченных реакций (исследования В.С.Хантера, Р.Йеркса, О.Л.Тинклпоу, Ф.Бойтендайка, В.Фишеля и др.). В Советском Союзе такие эксперименты проводились Н.Ю.Войтонисом, И.С.Беритовым, Г.З.Рогинским и др. Обычная схема проведения подобных опытов такова: на виду у животного прячется корм, который, однако, позволяется ему отыскать лишь некоторое время спустя. Обычно подопытное животное должно произвести выбор между несколькими местами, где может находиться спрятанная приманка. При успешном решении задачи имеет место реакция на отсутствующий стимул, что возможно лишь при наличии чувственных представлений.
Опыты по методу отсроченных реакций с успехом ставились над различными позвоночными, особенно же часто над обезьянами. Весьма эффектными были опыты, которые проводил Тинклпоу, когда на глазах у шимпанзе прятались фрукты (бананы), которые затем незаметно для животного подменялись значительно менее привлекательным кормом – салатом или капустой. Найдя спустя положенное время зелень; обезьяна медлила брать ее и продолжала, иногда с визгом, поиски. Последние продолжались до 33 секунд, в то время как в контрольном опыте, когда обезьяне с самого начала показывался салат, который затем не подменялся, поиски продолжались лишь 3 секунды и обезьяна спокойно съедала его. Напрашивается вывод, "то обезьяна ищет виденный ею вначале объект, руководствуясь зрительным представлением, которое, по Тинклпоу, отражает также определенные количественные и качественные показатели.
Войтонис на основании своих экспериментов также пришел к выводу, что обезьяны (макаки и шимпанзе) реагируют не на кормушку, содержащую приманку, а на корм как таковой, точнее, на специфический вид корма. То же самое относится и к собакам, но не к рыбам (опыты Бойтендайка и Фишеля).
Наличие у позвоночных (а возможно, и у некоторых высших беспозвоночных) представлений, выражающихся в отсроченных реакциях и способности к нахождению обходных путей (включая и явления экстраполяции), придает их поведению исключительную гибкость и намного повышает эффективность их действий на поисковых этапах поведенческих актов. Вместе с тем не следует переоценивать эти способности, ибо они не обязательно свидетельствуют о высоком уровне психического развития. Основываясь на развитой памяти, они встречаются у позвоночных на разных филогенетических ступенях в зависимости от образа жизни животного: наряду с животными, прячущими в разных местах запасы, эти способности свойственны хищникам, охотящимся на труднодосягаемых, прячущихся или лишь эпизодически появляющихся на виду животных.
На эту биологическую обусловленность реакции в отсутствии стимула, который до этого ее обусловливал, указал еще полвека тому назад советский зоолог и зоопсихолог Д.Н.Кашкаровна примере кошки, которая «стережет мышь и видит, что мышь появляется из норы. Мышь исчезает прежде, чем кошка начнет реагировать. Однако кошка встает и идет к норе. Здесь не может быть вопроса о том, что в момент реакции определяющий стимул отсутствует... Правильный выбор между тремя норами кошка могла бы сделать лишь на основании непосредственно предыдущего появления мыши, т.е. на основе представления о последней».* Что же касается обезьян, то, как видно из упомянутых исследований, мы явно имеем здесь дело с более сложными комплексными процессами.
* Кашкаров Д.Н. Современные успехи зоопсихологии. М.; Л., 1928. С. 123–124.
Способность высших позвоночных к предметному восприятию, способности к анализу и обобщению, а тем самым к формированию представлений являются важной предпосылкой образования сложных навыков, которые уже рассматривались раньше (см. ч. I) и которые составляют основное содержание накопления индивидуального опыта не только в сенсорной, но и в эффекторной сфере, точнее сказать, в моторно-сенсорной сфере, действующей как единая система.
Общение
У высших позвоночных особой сложности достигают и процессы общения. Как и у других животных, средства коммуникации включают у них элементы различной модальности – ольфакторные, тактильные.
Ольфакторная сигнализация, т.е. передача информации другим особям химическим путем, преобладает в территориальном поведении, особенно при маркировке местности у хищных и копытных. Для этого служат специальные железы, расположенные в различных участках кожи и выделяющие специфический пахучий секрет. Последний распространяется по воздуху или наносится на различные предметы (стволы и ветки растений, камни и т.д.). Пахучие метки производятся также с помощью испражнений. Выделениями пахучих желез маркируется и след, что способствует нахождению друг друга особями одного вида. Однако во всех этих случаях общение существенно не отличается от того, что мы наблюдаем, скажем, у насекомых.
Видотипичные, инстинктивные компоненты поведения позвоночных, служащие для акустического и оптического общения между животными, как правило, ритуализованы. Оптическое общение осуществляется прежде всего с помощью выразительных поз и телодвижений, о чем уже говорилось выше. Видоспецифические отличия выражаются часто в малозаметных человеку, но четко генетически фиксированных деталях, вызывающих совершенно специфические, зачастую ритуализованные реакции у особей того же вида. Особенно часто такие различия наблюдаются у близкородственных животных, причем прежде всего в сигнальных системах, связанных с размножением. Здесь они служат одним из факторов биологического разграничения видов.
Конкретные формы оптического общения отличаются у высших позвоночных большим разнообразием и дифференцированностью. Нередко они выражаются в специфических «диалогах» между двумя или несколькими особями, т.е. во взаимной демонстрации частей тела или поз. Особенно это относится к «ритуалам» запугивания или «импонирования». «Импонирование» выражает превосходство одной особи по отношению к другой, имеет и оттенок угрозы и вызова, но, в сущности, не носит агрессивного характера (в отличие от подлинного «запугивания», за которым часто следует нападение). Часто самцы «импонируют» самкам во время «ухаживания» за ними. Подчиненная особь отвечает на «импонирование» позой «покорности», чем окончательно исключается возможность столкновений между животными.
У млекопитающих оптическое общение часто сочетается с ольфакторным (рис. 43), так что выделение систем общения по отдельным модальностям является у этих животных в большой степени условным.
Это относится и к акустическому общению, в качестве примера которого следует, конечно, прежде всего напомнить о криках и песнях птиц. Хотя человеческое ухо неспособно улавливать все издаваемые птицами звуки (значительная часть их находится в ультразвуковом спектре ниже 50 кГц), мы поражаемся огромному разнообразию их голосовых реакций. К последним необходимо добавить и неголосовые звуки, как, например, свист крыльев при полете, постукивание дятла по стволу деревьев и т.д. Видоспецифичность таких звуков делает их пригодными для общения.

Рис. 43. Оптико-ольфакторное общение у грызунов (род Peromyscus). Стрелками обозначены места взаимного обнюхивания (по Эйзенбергу)
Для биологически адекватного реагирования на все эти звуковые сигналы существенной является видоспецифическая настройка слуховой системы на определенную часть спектра. Как показал советский исследователь биоакустики птиц В.Д.Ильичев, важнейшую роль играют в этой настройке периферические отделы слухового анализатора, выполняющие роль видотипических биологических фильтров. С их помощью осуществляется и специальная «подстройка» слуховой системы на особенно важные для особи звуки. Таким образом, здесь четко выступает характерная для всего инстинктивного поведения взаимообусловленность и единство врожденных функциональных (физиологических и этологических) и морфологических элементов.
Зоосемантика звуковых сигналов птиц охватывает практически все сферы их жизнедеятельности. Эти сигналы служат опознаванию особи (в частности, ее видовой принадлежности), оповещению других особей о физиологическом состоянии экспедиента (сюда относится, например, «попрошайничество») или об изменениях в окружающей среде (появление врага или, наоборот, кормового объекта и т.п.). При этом богатые зоопрагматические средства птицы позволяют экспедиенту передавать перцепиенту весьма детализованную информацию. Так, грач, подлетающий с кормом к гнезду, оповещает об этом насиживающую самку криком, могущим иметь четыре разных оттенка, на которые самка также отвечает разными звуками. Эти оттенки голосовых реакций птиц подчас настолько тонки, что не воспринимаются человеческим ухом.
Очень большое значение имеют не только четкие межвидовые различия акустического общения, но и индивидуальные, по которым особи узнают друг друга (самец – самку, птенцы – родителей и друг друга, члены стаи – друг друга и т.п.). Интересно, например, что, по данным В.Г.Торпа, в ряде случаев совместно поющие партнеры откликаются только на голос друг друга, но не реагируют на пение других особей. Особенно индивидуально различными являются территориальные крики птиц, оповещающих о занятости участка.
Большого разнообразия звуки достигают и у таких птиц, как куриные. Это обусловлено жизнью диких кур в труднопросматриваемых густых кустарниковых зарослях, где условия для оптического общения особенно неблагоприятны. Наиболее громкие и «впечатляющие» звуки куриных птиц – это крики петухов. Их слышимость (для человеческого уха) достигает 2 километров. В отличие, например, от звуков, с помощью которых петух подзывает кур к найденному им корму, кукареканье производится однократно, но зато дольше и на более высоких частотах. Это относится и к звуковым сигналам тревоги (например, при обнаружении врага), испуга или угрозы. Чем выше тональность, громкость и длительность этих сигналов, тем сильнее их эффект. Свое «кукареку» петух кричит на самых «высоких нотах», но всегда одинаковым образом: отклонения не превышают полтона.
Кукареканье – это своего рода вызов другим петухам, и те отвечают ответными криками, правда, лишь в том случае, если соперники – петухи одного «ранга». Если же в пределах видимости закричит петух, занимающий подчиненное положение, то господствующий попросту атакует его, «не удостаивая» ответным криком. Когда же перекликаются петухи, разделенные большим расстоянием и не видящие друг друга, то это типичные территориальные крики, т.е. акустическая маркировка местности, оповещение о занятости участков стайками кур – петухом и его «гаремом». Большое же биологическое значение крика заключается в том, что он предотвращает или хотя бы уменьшает частоту петушиных боев, а вместе с тем обеспечивает захват, освоение и защиту участков, а в конечном итоге расселение вида, но безвредным путем, ибо дело ограничивается одним запугиванием. Петух-соперник заранее предупреждается о том, что данный участок уже занят и ему придется искать другое пригодное для поселения место. И только в том случае, если все такие места уже заняты, начнутся бои за овладение участками. В этом случае победитель – животное более сильное не только в физическом, но и в психическом отношении – станет продолжателем рода, и это, конечно, тоже выгодно для процветания вида, для его прогрессивной эволюции. Аналогичную роль играет у других птиц пение. У домашних же кур крестьянский двор и прилегающие места остаются для петуха его участком, о занятии которого он непрестанно напоминает криком, хотя это поведение уже потеряло свое биологическое значение, поскольку человек обеспечивает все потребности домашних животных и управляет эволюцией вида. Надо думать, что чрезвычайная близость расположения участков домашних петухов заставляет их интенсивнее кукарекать, чем это имело бы место в дикой природе, так как в селе популяция кур «сверхплотная».
Мы не останавливаемся на звуковом общении млекопитающих, так как у них господствуют те же закономерности, что и у птиц.
Ригидность и пластичность в поведении высших позвоночных
Как уже указывалось, вопреки еще распространенному мнению инстинктивное поведение не теряет своей значимости в процессе эволюции, так как оно принципиально не может замещаться научением. Подчеркнем еще раз, что инстинктивное поведение является видовым поведением, научение – индивидуальным, и поэтому ничем не оправдано противопоставление этих двух основных категорий психической деятельности в качестве якобы различных генетических ступеней. На самом деле, как мы могли убедиться, прогресс инстинктивного поведения, в частности, в ряду позвоночных неразрывно связан с прогрессом индивидуально-изменчивого поведения, поэтому позвоночные с высокоразвитыми формами научения обладают и не менее развитыми сложными формами инстинктивного поведения.
Вместе с тем темпы эволюционных преобразований и специфическая роль в эволюции у инстинктивного поведения и научения различны. Главным образом, как показал Северцов, это относится к высшим позвоночным, особенно млекопитающим, у которых психика приобретает значение решающего фактора эволюции благодаря сильному развитию процессов научения, в том числе в высших их проявлениях – интеллектуальных действиях. Но при этом сохраняется в полной мере значение инстинктивной основы поведения, равно как медленно совершающихся изменений инстинктивного поведения. Приведенные примеры в достаточной мере показывают большое разнообразие истинных инстинктивных движений у высших позвоночных, вполне видотипичных, генетически фиксированных и достигающих в ряде случаев большой сложности. Конечно, и у высших позвоночных инстинктивные компоненты служат прежде всего для пространственно-временной ориентации наиболее жизненно важных поведенческих актов. Пространственная ориентация осуществляется и здесь на основе таксисов: тропо-, тело- и менотаксисов, т.е. типичных врожденных элементов поведения, к которым, однако, прибавляются мнемотаксисы, характеризующиеся индивидуальным запоминанием ориентиров. В последнем случае мы уже имеем дело с накоплением индивидуального опыта, и поскольку именно мнемотаксисы играют в жизни высших позвоночных особенно большую роль, то, следовательно, врожденные способы ориентации существенно обогащаются здесь индивидуально приобретаемыми способами.
В этой связи следует отметить, что, как и у других животных, биологическая адекватность реагирования на компоненты окружающей среды обеспечивается и у высших позвоночных именно инстинктивными элементами поведения. Биологически значимые объекты встречаются в окружающей животное среде в весьма разнообразных и, главное, постоянно меняющихся видах. Мы уже знаем, что их потребление или избегание, т.е. адекватное реагирование на биологические ситуации, возможно лишь в том случае, если животное руководствуется некими постоянными признаками этих объектов и ситуации. Именно это и происходит на генетически фиксированной, врожденной основе, когда животное реагирует на ключевые раздражители. К высшим позвоночным это относится точно так же, как к нижестоящим животным. Но у первых реакции на ключевые раздражители в очень большой степени конкретизируются индивидуальным опытом, дополнительно ориентируются хорошо развитым предметным восприятием. Тем самым инстинктивные действия приобретают и известное познавательное значение для животного, ибо помогают ему при ознакомлении с окружающей действительностью.
Особенно высокого уровня развития инстинктивное поведение достигает у высших позвоночных в ритуализованном общении животных друг с другом. Именно в сфере общения инстинктивные формы поведения достигают наибольшей стереотипности. Совершенно ясно, что иначе, без жестко фиксированных зоопрагматических средств, не может быть взаимопонимания между животными, т.е. не может быть подлинной передачи информации. Вместе с тем именно полноценное общение является необходимым условием для высшей интеграции в области поведения – интеграции поведения отдельных особей и целых сообществ.
Однако даже в сфере общения наличие и важное значение благоприобретаемых компонентов не вызывает сомнения. Так, у многих птиц птенцы не смогут петь видотипичным образом, если своевременно не слышали пения своих родителей. Несомненна роль научения и в образовании индивидуальных особенностей звукового общения, а также в многочисленных случаях акустического подражания у взрослых птиц. В последнем случае оказалось, что заимствованные звуки могут служить для общения наряду с собственными. На этой основе развивается межвидовое общение птиц, т.е. передача информации между особями разных видов. Здесь индивидуально приобретенные коммуникативные компоненты с четким сигнальным значением уже не только модифицируют и обогащают собственно инстинктивные компоненты, но и выступают во вполне самостоятельной роли.
Индивидуально приобретаемые элементы общения представляют особый «интерес, так как показывают возможность выхода за пределы закрытых коммуникативных систем животных. Особенно это относится к общению животных с человеком, в частности при дрессировке служебных собак и т.п. Вероятно, расширенное общение с животными сыграло в свое время немаловажную роль и при одомашнивании диких видов. Все это, разумеется, оказалось возможным лишь на высшем уровне перцептивной психики, т.е. у высших позвоночных. Если же иметь в виду предысторию антропогенеза, о чем пойдет речь ниже, то нельзя не признать, что способность высших позвоночных к расширению своих коммуникативных возможностей путем научения должна была стать важной предпосылкой зарождения человеческих форм общения.
Сказанное здесь о роли благоприобретаемых компонентов в инстинктивном поведении высших позвоночных вполне относится и к другим сферам поведения. Напомним, например, что у зубров и бизонов видотипичные признаки поведения появляются лишь на определенном этапе онтогенеза. Хотя здесь, несомненно, имеет место созревание некоторых видотипичных двигательных компонентов вне зависимости от частных внешних условий, все же и в этих процессах участвуют элементы научения. В остальном ограничимся здесь отсылкой к тому, что раньше говорилось по этому поводу об онтогенезе поведения.
Нет также надобности еще раз говорить о том, что в разных поведенческих актах удельный вес врожденных и благоприобретаемых компонентов различен, что и в навыках позвоночных содержатся качественно-гетерогенные элементы. Напомним лишь, что, не говоря уже о том, что навыки формируются на основе безусловнорефлекторных процессов, в их состав всегда входят консервативные двигательные компоненты, придающие навыкам в некоторых отношениях сходство с инстинктивными действиями. Более того, именно формирование таких консервативных компонентов составляет во многих случаях конечный итог и биологическую сущность образования навыка. Это . относится к двигательным стереотипам, автоматизмам, возникающим как результат закрепления навыка в ходе тренировки. Столь примитивные навыки встречаются не только у низших позвоночных. Наоборот, заученные автоматизированные действия играют немаловажную роль и в жизни высших млекопитающих, включая обезьян, а также человека (например, техника письма, повседневные «бытовые» движения и т.п.). У рыб же такими элементарными навыками, очевидно, ограничиваются их способности к научению. С этим связан и тот факт, что у рыб крайне трудно, если вообще возможно переделать положительное значение раздражителя на отрицательное и наоборот. Причиной тому является, в частности, недостаточное развитие ассоциативных функций головного мозга низших позвоночных. Как показал советский физиолог Б.Ф.Сергеев, только на уровне костистых рыб и земноводных появляется способность к образованию временных связей между всеми анализаторными системами и функциями организма, но еще отсутствуют внутри- и межанализаторные ассоциативные временные связи. Анализаторные системы еще разобщены, локализованы в различных отделах мозга.
Только у птиц и млекопитающих замыкательная функция головного мозга получает свое полное развитие, поэтому сложные навыки, в которых решающее значение имеют лабильные, гибкие компоненты, встречаются только у высших позвоночных. Они и определяют далеко идущую изменчивость, пластичность всего поведения животного, чему придавал столь большое адаптационное значение А.Н.Северцов. И действительно, сложные пластичные навыки в полной мере выполняют функцию быстрого приспособления организма к быстротечным изменениям среды. Пластичность навыков высшего порядка дополняет ригидность навыков низшего порядка и инстинктивных действий.
Эта пластичность проявляется в возможности быстрой перестройки навыка, в частности при превращении положительного или отрицательного раздражителя в противоположный. Другая важная особенность – возможность переноса навыка в новые условия, другими словами, адекватное использование накопленного чувственного и моторного опыта при существенных изменениях условий среды. Обеспечиваются эти возможности сильным развитием пластичности в сенсорной сфере, способностью к широким чувственным обобщениям, о чем уже шла речь выше.
Поясним сказанное на нескольких примерах. В первом эксперименте крыса получает приманку в результате ряда последовательных сложных инструментальных действий: она взбирается по лестнице на площадку, поднимает там эту лестницу с помощью перекинутой через блок бечевки, затем поднимается по лестнице на следующую площадку и там получает пищевое подкрепление. К этим фазам можно было бы добавить еще несколько – суть дела от этого не меняется: мы имеем здесь сложную операцию, в которой последовательность отдельных действий животного предопределена и строго фиксирована условиями задачи, т.е. теми компонентами среды (преградами), на которые по необходимости направлена активность животного.
В другом примере при относительно простой структуре операции выделяется способность к далеко идущему переносу операции в измененных условиях, что также весьма характерно для сложных навыков. В данном случае крыса, научившаяся находить кратчайший путь к «цели» в лабиринте, оказалась без какого-либо нового обучения способной к этому и после того, как лабиринт был поставлен вертикально. Поскольку животное при этом руководствуется и новыми ощущениями (с вестибулярного аппарата), то здесь с очевидностью имеют место и явления переноса в сенсорной сфере.
В третьем эксперименте крыса, освоившая сухопутный лабиринт, тут же с неменьшим успехом проплыла этот лабиринт после того, как он был заполнен водой. Поскольку при плавании производятся иные движения, чем при ходьбе, мы можем здесь констатировать далеко идущий перенос в моторной сфере при сохранении прежней ориентации в пространстве.
Даже в первом примере, а тем более в остальных, невозможно расценивать поведение подопытного животного как простую цепь механически усвоенных движений, ибо в таком случае совершенно исключалась бы возможность переноса операции и сохранения навыка в измененных условиях. Сложные навыки представляют собой исключительно динамические моторнорецепторные системы, обеспечивающие на основе высокоразвитой ориентировочной деятельности выработку весьма пластичных двигательных программ. Процесс ориентировки сливается здесь с двигательной активностью, а нахождение верного решения задачи формируется в ходе этой активности на основе высокоразвитого чувственного обобщения.
Отмеченными качествами сложные навыки выделяются среди других видов научения, и именно благодаря им сложные навыки стали предпосылками и основой развития высших форм психической деятельности животных – интеллектуальных действий.
Проблема интеллекта животных
Предпосылки и элементы интеллектуального поведения животных
Интеллектуальное поведение является вершиной психического развития животных. Однако, говоря об интеллекте, «уме» животных, их мышлении необходимо прежде всего отметить, что чрезвычайно трудно точно указать, по поводу каких животных можно говорить об интеллектуальном поведении, а по поводу каких – нет. Очевидно, речь может идти лишь о высших позвоночных, но явно не только о приматах, как это до недавнего времени принималось. Вместе с тем интеллектуальное поведение животных является не чем-то обособленным, из ряда вон выходящим, а лишь одним из проявлений единой психической деятельности с ее врожденными и благоприобретаемыми аспектами. Интеллектуальное поведение не только теснейшим образом связано с разными формами инстинктивного поведения и научения, но и само складывается (на врожденной основе) из индивидуально-изменчивых компонентов поведения. Оно является высшим итогом и проявлением индивидуального накопления опыта, особой категорией научения с присущими ей качественными особенностями. Поэтому интеллектуальное поведение дает наибольший приспособительный эффект, на что и обратил особое внимание А.Н.Северцов, показав решающее значение высших психических способностей для выживания особей и продолжения рода при резких, быстро протекающих изменениях в среде обитания.
Предпосылкой и основой развития интеллекта животных – во всяком случае в направлении, ведущем к человеческому сознанию – является манипулирование, причем прежде всего с биологически «нейтральными» объектами. Особенно, как уже было показано, это относится к обезьянам, для которых манипулирование служит источником наиболее полных сведений о свойствах и структуре предметных компонентов среды, ибо в ходе манипулирования происходит наиболее глубокое и всестороннее ознакомление с новыми предметами или новыми свойствами уже знакомых животному объектов. В ходе манипулирования, особенно при выполнении сложных манипуляций, происходит обобщение опыта деятельности животного, формируются обобщенные знания о предметных компонентах окружающей среды, и именно этот обобщенный двигательно-сенсорный опыт составляет главнейшую основу интеллекта обезьян.
О манипулировании человекообразных обезьян «биологически индифферентными» предметами Павлов говорил: «Это же – самая настойчивая любознательность. Так что нелепое утверждение, будто у животных ее нет, нет в зачатке того, что есть у нас и что в конечном счете создало науку, – не отвечает действительности».* В качестве примера Павлов сослался на наблюдавшееся им у шимпанзе манипулирование предметами, в частности коробкой, в которой нет «никаких апельсинов, ни яблок». Тем не менее обезьяна «долгое время возится... над решением механических задач, которое не обещает ей никаких выгод, никакого материального удовлетворения».**
* Павловские среды. Т. II. М.; Л., 1949. С. 166.
** Там же.
Эта, по Павлову, «чистейшая, бескорыстная любознательность» и заставляет обезьяну изучать объект манипулирования в ходе активного воздействия на него. При этом одновременно и во взаимодействии друг с другом включаются в познавательную деятельность животного разные сенсорные и эффекторные системы. Ведь манипулирующая обезьяна почти непрерывно следит за движениями своих рук; под пристальным зрительным контролем производятся самые разнообразные действия как без разрушения целостности объекта: поворачивание в разные стороны, облизывание, поглаживание, придавливание, перекатывание и т.п., так и деструктивного порядка: разламывание, разрывание, вычленение отдельных деталей и т.д. (рис. 24).
Особую познавательную ценность представляют деструктивные действия, так как они позволяют получить сведения о внутренней структуре предметов. При манипулировании животное получает информацию одновременно по ряду сенсорных каналов, но преобладающее значение имеет у обезьян сочетание кожно-мышечной чувствительности рук со зрительными ощущениями. Кроме того, в обследовании объекта манипулирования участвуют также обоняние, вкус, тактильная чувствительность околоротовых вибрисс, иногда слух и т.д. Эти виды чувствительности сочетаются с кожно-мышечной чувствительностью эффекторов (ротового аппарата, передних конечностей) кроме обезьян и у других млекопитающих, когда те манипулируют предметами. В итоге животные получают комплексную информацию об объекте как едином целом и обладающем разнокачественньми свойствами. Именно в этом и заключается значение манипулирования как основы интеллектуального поведения.
Необходимо, однако, подчеркнуть, что первостепенное значение имеют для интеллектуального поведения зрительные восприятия и особенно зрительные обобщения, о которых уже раньше шла речь. Насколько развита способность к формированию обобщенных зрительных образов даже у крыс – показывает следующий эксперимент, в котором крысы с успехом решали очень трудную задачу: животному надо выбрать из трех предъявленных фигур (вертикальные и горизонтальные полосы) одну несходную по сравнению с двумя другими. Местонахождение и рисунок такой фигуры постоянно меняются, следовательно, это будут то вертикальные, то горизонтальные полосы, расположенные то слева, то справа, то посередине (в неправильной последовательности). Таким образом, подопытное животное могло ориентироваться лишь по одному, крайне обобщенному признаку – несхожести одного рисунка по сравнению с остальными. Мы имеем здесь, таким образом, дело со зрительным обобщением, близким к абстрагированию, свойственному мыслительным процессам.
С другим элементом интеллектуального поведения, на этот раз в двигательной сфере, мы встречались при описании опытов с «проблемными ящиками». И здесь мы имеем дело со сложными многофазовыми навыками, поскольку у высших млекопитающих, например енотов, можно сравнительно легко добиться решения задач, при которых животному приходится в определенной последовательности открывать совокупность разнообразных запирающих приспособлений. Как и при описанном опыте с подтягиванием крысой лестницы, енот может решить такую задачу лишь при соблюдении строго определенной последовательности действий. Но разница заключается в том, что в отличие от крысы еноту приходится самому находить эту последовательность, и это поднимает его деятельность, безусловно, на более высокий уровень. Правда, по некоторым данным и крысы способны на это.
Необходимо, однако, подчеркнуть, что даже высшие позвоночные решают инструментальные задачи труднее, чем локомоторные. Л. Кардош отметил в этой связи, что в психической деятельности животных преобладает познавание пространственных отношений (см. ч. I, гл. 3), постигаемых ими с помощью локомоторных действий. У обезьян же, особенно человекообразных, локомоторное познавание пространственных отношений теряет свою доминирующую роль за счет сильного развития манипуляционных действий. Однако только человек может полностью освободиться от направляющего воздействия пространственных отношений, если этого требует познание временно-причинных связей.
Решение многофазовых инструментальных задач у обезьян изучалось рядом исследователей, в частности Н. Н. Ладыгиной-Котс. В своей монографии «Приспособительные моторные навыки макака в условиях эксперимента» она подвела итог многочисленным опытам, в которых использовались весьма разнообразные сочетания запирающих механизмов. Эти эксперименты показали, что низшая обезьяна (макак-резус) способна научиться отмыкать большие серии запирающих механизмов, хотя лучше справляется с одиночными установками. Характерными явились при этом многочисленные и многообразные нащупывающие движения рук, «экспериментирование». Ввиду ее большой торопливости наиболее трудными оказались для обезьяны те приспособления, которые не отмыкались легкими быстрыми движениями. Легче всего производились такие движения, как вытягивание, отведение, притягивание, опускание и др., труднее всего – отодвигание и вращение, вывертывание. Вообще в поисках пунктов задержки и пути преодоления препятствий преобладающая роль принадлежала кинестетическим, а не зрительным восприятиям. Интересно, что по многим из этих особенностей, как показали более поздние исследования, действия низших обезьян в опытах с запирающими механизмами напоминают таковые енотов.
Чрезвычайно важной предпосылкой интеллектуального поведения является и способность к широкому переносу навыков в новые ситуации. Эта способность вполне развита у высших позвоночных, хотя и проявляется у разных животных в разной степени. В. П. Протопопов приводит следующий пример переноса приобретенного опыта в новую ситуацию у собаки. Первоначально подопытная собака научилась открывать нажатием лапы щеколду на дверце «проблемной клетки», в которой находилась приманка. В других опытах та же собака научилась затем подтягивать зубами и лапами кусок мяса за веревку, которая лежала перед ней на полу. После этого была создана третья ситуация, содержавшая элементы первых двух: на клетке, применявшейся в первой ситуации, щеколда была поднята на такую высоту, что собака не могла ее достать лапой, но к щеколде привязывалась веревка, потянув за которую можно было ее открыть. Когда собака была подведена к клетке, она сразу же, без всяких других проб, схватила зубами веревку и, потянув, открыла щеколду. Таким образом, задача была немедленно решена в новой ситуации, несмотря на то что прежние элементы располагались в ней совсем по-иному: веревка висела, а не лежала горизонтально на полу, на конце ее было привязано не мясо, а щеколда, которая к тому же находилась на другом месте – наверху. К тому же щеколда отпиралась в первых опытах движением лапы, а затем – с помощью зубов. «...Новый навык, – пишет по этому поводу Протопопов, – вырабатывается сразу, "внезапно", но эта внезапность... обусловлена вполне определенными следами прошлого опыта, которые под влиянием стимула вступают путем как бы взрывчатого замыкания в новую временную связь, и создается новая нервная структура и новая реакция, отличающаяся от прежних двух и в рецепторной и эффекторной части... Подобные навыки... могут по своему внешнему проявлению имитировать разумное поведение и, если не знать этапов их возникновения, можно прийти к ошибочным антропоморфическим заключениям».*
* Протопопов В.П. Исследование высшей нервной деятельности в естественном эксперименте. Киев, 1950. С. 21–23.
К подобным антропоморфическим выводам пришел, например, Н. Р. Ф. Майер, признавший крыс способными «рассуждать». Обоснованием этого заключения послужили результаты его экспериментов по выработке у крыс отсроченных реакций, в ходе которых эти животные оказались в состоянии связывать элементы прежнего опыта, которые раньше никогда не сочетались в их поведении. Как было показано, это происходило и в опытах Протопопова с собакой.
Итак, способности высших позвоночных к разнообразному манипулированию, к широкому чувственному (зрительному) обобщению, к решению сложных задач и переносу сложных навыков в новые ситуации, к полноценной ориентации и адекватному реагированию в новой обстановке на основе прежнего опыта являются важнейшими элементами интеллекта животных. И все же сами по себе эти качества еще недостаточны, чтобы служить критериями интеллекта, мышления животных. Тем более, как указывалось, невозможно признать такими критериями, например, высокоразвитые способности к оптическому обобщению у пчел.
Критерий интеллектуального поведения животных
Отличительная особенность интеллекта животных заключаются в том, что в дополнение к отражению отдельных вещей возникает отражение их отношений и связей (ситуаций). Отчасти это имеет, конечно, место и при некоторых сложных навыках, что лишний раз характеризует последние как переходную форму к интеллектуальному поведению животных. Это отражение происходит в процессе деятельности, которая по своей структуре, согласно Леонтьеву, является двухфазной.
Мы уже видели, что сложные навыки животных большей частью являются многофазными. Однако эти фазы, будь это вскарабкивание крысы с площадки на площадку с помощью подтягиваемой лестницы или последовательное отмыкание затворов «проблемного ящика», по существу своему являются лишь цепью, суммой однозначных равнокачественных этапов последовательного решения задачи. По мере же развития интеллектуальных форм поведения фазы решения задачи приобретают четкую разнокачественность: прежде слитая в единый процесс деятельность дифференцируется на фазу подготовления и фазу осуществления. Именно фаза подготовления составляет характерную черту интеллектуального поведения. Как указывает Леонтьев, интеллект возникает впервые там, где возникает процесс подготовки возможности осуществить ту или иную операцию или навык.
В конкретных экспериментальных исследованиях двухфазность интеллектуальных действий проявляется, например, в том, что обезьяна достает сперва палку, чтобы затем с помощью этой палки сбить высоко подвешенный плод, как это имело место в широко известных опытах немецкого психолога В. Кёлера. В других экспериментах обезьяна могла овладеть приманкой лишь в том случае, если сперва оттолкнет ее от себя палкой к такому месту, где ее (после обходного движения) можно достать рукой (рис. 44).
Производилось и много других экспериментов, в которых обезьяны должны были решить задачу с употреблением орудия (чаще всего палки). Так, в опытах Г.З.Рогинского шимпанзе, имевшие опыт манипулирования палками, сразу употребляли таковые для доставания приманки. Но низшие обезьяны, кроме одной (павиан чакма), к этому оказались не сразу способны. Все же Рогинский отвергает мнение В.Кёлера о наличии разрыва между психикой человекообразных и низших обезьян.
Рис. 44. Схема сложной задачи, для решения которой обезьяна должна привязанной к дереву палкой оттолкнуть плод в ящике через щель к противоположной (решетчатой) стенке, а затем обойти ящик. Прикорм (и) первоначально виден как через решетку, так и через щель в стенке, но не может непосредственно браться рукой (опыт Кёлера)
Советский зоопсихолог Л. С. Новоселова сумела своими исследованиями выявить генезис употребления палок при решении сложных задач у шимпанзе. Она показала, что употребление палки формируется как индивидуально-приспособительное действие, но не является врожденной формой поведения. При этом намечается несколько этапов – от оперирования всей рукой как рычагом к специализированным действиям кистью, которая уже не только удерживает палку, но и направляет ее движения в соответствии со специфическими свойствами орудия.
Н. Н. Ладыгина-Котс детально изучала у шимпанзе процесс подготовки и даже изготовления-орудия, необходимого для решения технически несложной задачи – выталкивания приманки из узкой трубки. На глазах у шимпанзе в трубку закладывалась приманка таким образом, что ее нельзя было достать просто пальцами. Одновременно с трубкой животному давались различные предметы, пригодные для выталкивания прикорма после некоторой их «доработки» (рис. 45). Подопытная обезьяна вполне (хотя и не всегда немедленно) справлялась со всеми этими задачами.
Рис. 45. Экспериментальное изучение орудийной деятельности у шимпанзе (опыты Ладыгиной-Котс). Некоторые предметы, предъявленные обезьяне (ветка, палка, обмотанная веревкой, изогнутая и спиралевидно закрученная проволока, проволочная сетка, кусок плетеной корзины)
В этих опытах также четко выступает двухфазность интеллектуального действия: приготовление орудия – первая, подготовительная фаза, доставание приманки с помощью орудия – вторая фаза. Первая фаза вне связи со следующей фазой лишена какого бы то ни было биологического смысла. Вторая фаза – фаза осуществления деятельности – в целом направлена на удовлетворение определенной биологической потребности животного (в описанных опытах – пищевой).
По Леонтьеву, первая, подготовительная фаза побуждается не самим предметом (например, палкой), на который она направлена, а объективным отношением палки к приманке. Реакция на это отношение и есть подготовка второй фазы, фазы осуществления, которая направлена на объект («цель»), побуждающий всю деятельность животного. Вторая фаза включает, таким образом, в себя определенную операцию, закрепляемую в виде навыка.
Большое значение как один из критериев интеллектуального поведения имеет и то обстоятельство, что при решении задачи животное пользуется не одним стереотипно выполняемым способом, а пробует разные способы, которые являются результатом ранее накопленного опыта. Следовательно, вместо проб различных движений, как это имеет место при неинтеллектуальных действиях, при интеллектуальном поведении имеют место пробы различных операций, что позволяет решить одну и ту же задачу различными способами. Перенос и, пробы различных операций при решении сложной задачи находят у обезьян свое выражение, в частности, в том, что они практически никогда не пользуются орудиями совершенно одинаковым образом.
Таким образом, при интеллектуальном поведении мы имеем дело с переносом операции, причем этот перенос не требует, чтобы новая задача была непосредственно сходной с прежней. Операция, как отмечает Леонтьев, перестает быть неподвижно связанной с деятельностью, отвечающей определенной задаче. И здесь мы можем проследить преемственность от сложных навыков.
Поскольку интеллектуальное поведение животных характеризуется отражением не просто предметных компонентов среды, а отношений между ними, здесь осуществляется и перенос операции не только по принципу сходства вещей (например, преград), с которыми была связана данная операция, но и по принципу сходства отношений, связей вещей, которым она отвечает.
Формы мышления
На основе многолетних экспериментальных исследований Ладыгина-Котс пришла к выводу, что мышление животных имеет всегда конкретный чувственно-двигательный характер, что это мышление в действии, причем эти действия являются всегда предметно-отнесенными. Такое мышление в действиях, по Ладыгиной-Котс, представляет собой практический анализ и синтез, который при орудийной деятельности осуществляется в ходе непосредственного обращения с объектами, в ходе их обследования, обработки и применения. Но при этом не исключаются, а, наоборот, играют большую роль те обобщенные зрительные представления, о которых шла речь выше.
В соответствии с этим Ладыгина-Котс считает возможным выделить две различные по сложности и глубине формы мышления (правда, только у человекообразных обезьян).
Первая форма характеризуется установлением связей раздражителей (предметов или явлений), непосредственно воспринимаемых животным в ходе его деятельности. Это анализ и синтез в наглядно обозреваемой ситуации. Примером может служить выбор обезьяной предметов, пригодных для употребления в качестве орудия, с учетом их величины, плотности, формы и т.д.
Вторая форма характеризуется установлением связей между непосредственно воспринимаемыми раздражителями и представлениями (зрительными следами). Так, в описанных опытах, в которых шимпанзе приходилось выталкивать лакомство из трубы с помощью орудия, мыслительные операции такого типа проявлялись в изготовлении орудия, например в распутывании клубка проволоки и ее выпрямлении. Особенно убедительным доказательством существования этой высшей формы мышления является опыт, при котором обезьяне вместе с трубой давалась дощечка значительно более широкая, чем диаметр трубы. Шимпанзе оказался в состоянии совершенно самостоятельно отчленять от доски узкие лучины и использовать их в качестве орудия для выталкивания приманки из трубы.
Подобное поведение обезьяны можно здесь объяснить только тем, что у нее в ходе предыдущей деятельности сформировалось обобщенное зрительное представление о предмете типа палки (но только в ситуации данной задачи). Решающая роль предшествующего опыта в формировании подобных зрительных "представлений достаточно четко выступает в этой же серии экспериментов. Ведь еще до решения описанной задачи обезьяна накапливала опыт по «доработке» весьма различных предметов, среди которых были и фигурные дощечки (рис. 46), превращение которых в пригодные для решения задачи орудия было для шимпанзе делом не более трудным, чем обламывание боковых побегов ветки (рис. 45). Вместе с тем изготовление орудия из такой фигурной дощечки требует отщепления части ее в продольном направлении. Подобные действия и накапливаемый при этом опыт и подготовили обезьяну к решению задачи с широкой дощечкой, которая ей давалась в одном из следующих экспериментов.
На основе установления связи обобщенного зрительного представления о необходимом предмете (типа палки) с непосредственным восприятием второго предмета (трубы), также данного в конкретной ситуации опыта, обезьяна сумела вычленить (по воображаемой линии!) часть из целого – лучину из доски, причем таким образом, что она оказалась пригодной служить орудием для выталкивания приманки.
Рис. 46. Фигурные доски с расширениями на концах или в середине, предлагавшиеся шимпанзе в опытах Ладыгиной-Котс.
Таким образом, шимпанзе в состоянии мысленно разлагать целые объекты на детали, равно как и сложные фигуры – на их составные части. Как уже указывалось, ведущую роль в поведении, и в частности в интеллектуальных действиях обезьян играют их руки, тактильно-кинестетическая чувствительность руки. И. П. Павлов говорил поэтому с полным основанием о «ручном мышлении» обезьян. Сочетание тактильно-кинестетической чувствительности со зрением дает обезьяне большие преимущества для установления пространственно-временных связей для практического анализа и синтеза. Именно это существенное расширение и углубление сенсорной сферы легло у обезьян в основу того, что И. П. Павлов обозначил как «улавливание постоянной связи между вещами» (или «нормальной связи вещей»).*
* Павловские среды. Т. III. С. 262.
Биологическая ограниченность интеллекта животных
Наряду со всем этим надо четко представить себе биологическую ограниченность интеллектуального поведения обезьян. Как и все другие формы поведения, оно всецело определяется образом жизни и чисто биологическими закономерностями, рамки которых не может перешагнуть даже самая умная обезьяна. Так, например, шимпанзе на воле каждый вечер устраивают себе искусно переплетенные спальные гнезда из ветвей и листьев, но, по свидетельству английской исследовательницы поведения обезьян Дж. ван Лавик-Гудолл, они никогда не сооружают навесов и остаются совершенно беззащитными под проливным тропическим дождем.
Очень редко обезьяны пользуются на воле и орудиями. Правда, имеются отдельные наблюдения об орудийных действиях шимпанзе при добывании пищи или нападении. Но, как и другие человекообразные обезьяны, шимпанзе вполне обходятся в повседневной жизни без орудий. С другой стороны, другие позвоночные (каланы, галапагосские дятловые вьюрки и др.) систематически пользуются предметами в качестве орудий. Уже это указывает на то, что орудийные действия сами по себе не обязательно являются критериями высокоразвитой психической деятельности животных.
Биологическую ограниченность интеллекта антропоидов вскрывает и анализ экспериментальных данных. Так, Ладыгина-Котс показала, что зрительные образы, представления человекообразных обезьян являются значительно более слабыми, чем у человека, и всегда связаны с компонентами окружающей обстановки (ситуационная связанность представлений).
Эта ограниченность интеллектуального поведения неоднократно проявлялась в опытах Ладыгиной-Котс, когда шимпанзе допускал «нелепые» ошибки при употреблении предметов, предоставленных ему для выталкивания приманки из трубы. Так, например, он пытался втолкнуть в трубу кусок фанеры вопреки явному несоответствию ее ширины и принимался обгрызать его лишь после ряда таких неудачных попыток. Подчас неадекватность действий обусловливалась преобладанием деструктивного манипулирования (рис. 47). Кроме того, была проведена специальная серия опытов, в которых обезьяне давалась труба, закрытая с одного конца, и крючок. Если приманка, заложенная в такую трубу, была прикреплена к нити, шимпанзе легко вытягивал ее оттуда. Несмотря на это обезьяна не сумела адекватно применить крючок и, более того, чаще всего отламывала загнутую его часть как мешавший элемент. Ладыгина-Котс писала по этому поводу, что «шимпанзе оказался неспособным перейти от шаблонного привычного приема выталкивания приманки прямым, гладким единичным орудием к использованию приема притягивания к себе крюком», и усматривала в этом «недостаточную пластичность психики шимпанзе, ограниченность его мышления».*
* Ладыгина-Котс Н.Н. Конструктивная и орудийная деятельность высших обезьян (шимпанзе). М., 1959. С. 247.
Шимпанзе, по Ладыгиной-Котс, «не в состоянии схватить сразу существенные особенности в новой ситуации и установить новые связи на основе осмысливания непосредственно воспринимаемых отношений между предметами».*
* Там же.
Рис. 47. Предметы, предлагавшиеся шимпанзе, при использовании которых обнаруживалась ограниченность интеллектуального поведения этой обезьяны: палочки пригодны для выталкивания приманки из трубы только в связанном виде, обезьяна же развязала их и пыталась применить поодиночке; крестовидно соединенные планки надо было соединить вместе, сдвинутые же под углом – распрямить до прямой, как это показано на нижней части рисунка. Вместо этого шимпанзе прежде всего разъединил планки и пытался применить их по отдельности (опыты Ладыгйной-Котс)
Этот вывод Ладыгиной-Котс подтверждается экспериментами и других исследователей. Так, шимпанзе показал ситуационную связанность своих представлений и неспособность охватить существенное изменение в прежней ситуации в следующем опыте: шимпанзе предлагается с помощью палки подкатывать яблоко к клетке вокруг невысокой стенки. После овладения обезьяной этим навыком убирается часть стенки непосредственно перед клеткой, в результате чего было бы удобнее непосредственно притягивать яблоко палкой. Тем не менее обезьяна продолжает выполнять прежнее сложное, трудное действие, отталкивая яблоко от себя и обводя его вокруг стенки (опыт Э. Г. Вацуро, рис. 48).
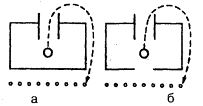
Рис. 48. Опыт Вацуро, вскрывающий ограниченность и качественное своеобразие интеллекта человекообразных обезьян. Пояснения см в тексте
Даже самые сложные проявления интеллекта обезьян представляют собой в конце концов не что иное, как применение в новых условиях филогенетически выработанного способа действия. Ведь давно уже подмечалось сходство притягивания приманки с помощью палки с притягиванием плода, растущего на ветке. Войтонис и Ладыгина-Котс указывали на то, что развитая способность обезьян к практическому анализу связана с особенностями их питания; высокоразвитые сенсомоторные функции руки, их сочетание со зрением и как следствие отличительные познавательные способности обезьян Фабри объясняет функциональными особенностями их хватательных способностей (см. гл. 3) и т.д. Эта биологическая обусловленность всей психической деятельности обезьян, включая и антропоидов, и является причиной отмеченной ограниченности их интеллектуальных способностей, причиной их неспособности к установлению мысленной связи между одними лишь представлениями и их комбинированием в образы. Неспособность же мысленно оперировать одними лишь представлениями с неизбежностью приводит к неспособности понимать результаты своих действий, понимать истинные причинно-следственные связи. Это возможно лишь с помощью понятий, которые у обезьян, как и у всех других животных, по указанным причинам всецело отсутствуют.
В заключение приходится признать, что проблема интеллекта животных еще совершенно недостаточно изучена. По существу до сих пор проведены обстоятельные экспериментальные исследования только над обезьянами, преимущественно высшими, в то время как о возможности интеллектуальных действий у других позвоночных еще почти нет доказательных экспериментальных данных. Вместе с тем, как уже говорилось, сомнительно, чтобы интеллект был присущ только приматам.
Глава 3 ЭВОЛЮЦИЯ ПСИХИКИ И АНТРОПОГЕНЕЗ
С древнейших времен человек пытался как-то осознать свое отношение к миру животных, искал признаки сходства и различия в поведении последних. Об этом свидетельствует та исключительная роль, которую играет поведение животных в бесчисленных древних сказках, легендах, верованиях, культовых обрядах и т.п.
С зарождением научного мышления проблема «души» животного, его психики и поведения стала важной составной частью всех философских концепций. Как уже отмечалось при обсуждении проблемы инстинкта и научения, часть древних мыслителей держалась мнения о близком родстве и одинаково высоком уровне психической жизни человека и животных, другие же, наоборот, отстаивали превосходство человеческой психики, а иные категорически отрицали всякую ее связь с психической деятельностью животных. Как и в других областях знания, теоретические построения античных ученых, лишь отчасти основанные на непосредственных наблюдениях, определяли затем на протяжении тысячелетия и более взгляды на поведение животных, если не считать ненаучных клерикально-догматических толкований средневековья.
Предшественники творцов эволюционного учения отказались от произвольных толкований о сходстве и различиях психики животных и человека и обратились к фактам, которые добывало зарождающееся научное естествознание. Так, уже Ж. О. Ламеттри обратил внимание на сходство строения мозга человека и млекопитающих. При этом он отметил, что у человека значительно больше мозговых извилин.
В борьбе за эволюционное учение, стремясь обосновать положение о непрерывности развития всего органического мира как единого целого, Ч. Дарвин и его последователи односторонне подчеркивали сходство и родство всех психических явлений начиная от низших организмов до человека. Особенно это относится к отрицанию Дарвином качественных различий между психикой человека и психикой животных. Как бы приближая человека к животным и «поднимая» до него последних, Дарвин приписывал животным человеческие мысли, чувства, воображения и т.д.
Одностороннее понимание генетического родства психики человека и животных было- как уже отмечалось, подвергнуто критике В.А Вагнером. Вагнер подчеркивал, что сравнение психических функций животных надо проводить не с психикой человека, а с психикой форм, непосредственно предшествующих данной группе животных и за ней следующих. При этом он указывал на наличие общих законов эволюции психики, без познания которых невозможно понимание человеческого сознания. Такой эволюционный подход позволяет достоверно выявить предысторию антропогенеза, и в частности биологические предпосылки зарождения человеческой психики.
При этом, однако, необходимо учесть, что судить о происхождении человеческого сознания, как и о процессе антропогенеза вообще, можно лишь косвенно, лишь по аналогии с тем, что наблюдаем у ныне живущих животных. Эти животные, однако, прошли длительный путь адаптивной эволюции, и их поведение носит глубокий отпечаток специализации к особым условиям их существования.
Если, к примеру, взять высших позвоночных, к которым в биологическом отношении относится и человек, то здесь наблюдается ряд боковых ответвлений в эволюции психики, не имеющих отношения к линии, ведущей к антропогенезу, а отражающих лишь специфическую биологическую специализацию отдельных групп животных. Наиболее яркий пример представляют собой в этом отношении птицы. То же относится и к млекопитающим, отдельные отряды которых воплощают подобную специализацию к специфическому образу жизни. И даже в отношении приматов мы должны считаться с той же специализацией, причем ныне живущие антропоиды в ходе своего развития от вымерших общих с человеком предков не только не приблизились к человеку, а, наоборот, отдалились. Поэтому они, судя по всему, находятся сейчас на более низком психическом уровне, чем этот предок.
Отсюда следует также, что все, даже наиболее сложные, психические способности обезьян всецело определяются условиями их жизни в естественной среде, их биологией, а с другой стороны, служат только приспособлению к этим условиям. Особенности образа жизни обусловливают специфические особенности психических процессов, в том числе и мышления обезьян.
Обо всем этом важно помнить, когда речь идет о поиске биологических корней и предпосылок зарождения человеческого сознания. По поведению ныне существующих обезьян и других животных мы можем судить лишь об уровнях и направлениях психического развития, ведших к человеку, и об общих закономерностях этого процесса.
Проблема происхождения трудовой деятельности
Эволюция гаптических и сенсорных функций высших млекопитающих
Общеизвестно, что решающий фактор превращения животного предка – ископаемой человекообразной обезьяны - в человека был открыт около ста лет тому назад Ф. Энгельсом: труд, создавший человека, создал и человеческое сознание. Трудовая деятельность, членораздельная речь, а на их основе и общественная жизнь определяли развитие человеческой психики и, таким образом, являются отличительными критериями психической деятельности человека по сравнению с таковой животных. Поэтому для выяснения конкретных условий возникновения сознания необходимо отыскать в мире животных возможные биологические предпосылки этих форм человеческой деятельности и проследить вероятный путь их развития.
Труд с самого своего зарождения был ручным, таким же по необходимости было и изначальное человеческое мышление. У современных обезьян, как уже отмечалось, оно так и осталось чисто «ручным мышлением». Рука – орган и продукт человеческого труда: развившись из руки обезьяны, став органом труда, человеческая рука достигла благодаря труду, причем только благодаря труду, как подчеркивал Энгельс, «той высокой ступени совершенства, на которой она смогла, как бы силой волшебства, вызвать к жизни картины Рафаэля, статуи Торвальдсена, музыку Паганини».*
* Энгельс Ф. Диалектика природы. М., 1949. С. 133.
Таким образом, рука (ее развитие и качественные преобразования) занимает центральное место в антропогенезе как в физическом, так и в психическом отношении. При этом основную роль сыграли ее исключительные хватательные (гаптические) способности (рис. 49). Биологические предпосылки зарождения трудовой деятельности необходимо, следовательно, искать прежде всего в особенностях хватательной функции передних конечностей млекопитающих.
Здесь мы сталкиваемся с весьма интересным вопросом: почему, собственно, именно обезьяны стали предками человека? Почему начало развитию разумных существ не могла дать другая группа млекопитающих, тем более что хватательная функция не является привилегией только обезьян. В поисках ответа на эти вопросы Фабри изучал в сравнительном аспекте у обезьяны и других млекопитающих взаимоотношения между главной (локомоторной) и дополнительными (манипуляционными) функциями передних конечностей и установил, что решающее значение имели для процесса антропогенеза антагонистические отношения между главной и дополнительными функциями передних конечностей. Активное участие одновременно обеих передних конечностей в обращении с предметами в манипулировании связано с частым их освобождением от функций опоры и передвижения, что препятствует специализации к длительному быстрому бегу.
Рис. 49. Гаптические движения рук обезьян (по Даниловой)
Вместе с тем при редукции манипуляционных функций страдают прежде всего хватательные способности конечностей. Как уже раньше указывалось, часть дополнительных функций передних конечностей переходит при этом к ротовому аппарату. В наименьшей степени дополнительные, и в том числе хватательные, функции подавлены у медведей, енотов и некоторых других млекопитающих (преимущественно хищных и грызунов), но и здесь эволюция двигательной активности определялась антагонизмом между главной и дополнительными функциями их передних конечностей.
Единственное исключение среди млекопитающих, по Фабри, составляют приматы. Основная, и главное, первичная форма их передвижения состоит в лазанье путем хватания веток, и эта форма локомоции составляет, таким образом, основную функцию их конечностей. Но такой способ передвижения сопряжен с усилением подвижности пальцев и сохранением противопоставления первого пальца остальным, что гармонично сочетается с требованиями, предъявляемыми манипулированием предметами. Поэтому у обезьян, и притом только у них, главная и дополнительная функции передних конечностей не находится в антагонистических отношениях, а гармонично сочетаются друг с другом.
Это обстоятельство составляет одно из наиболее существенных отличий эволюции двигательной активности приматов. На основе гармоничного сочетания манипуляционных функций с локомоцией и их взаимного усиления и стало возможным столь мощное развитие тех исключительных двигательных возможностей, которые возвысили обезьян над остальными млекопитающими и заложили основу формирования специфических двигательных способностей человеческой руки.
Как показали далее исследования Фабри, благодаря отсутствию антагонизма между функциями передней конечности приматов у них опять-таки в отличие от других млекопитающих гаптическая функция руки развивалась одновременно по двум направлениям: 1) в сторону увеличения полного обхватывания предметов и 2) в сторону увеличения гибкости, вариабельности хватательных движений. А ведь только такое развитие могло служить достаточной эволюционный основой для зарождения употребления орудий труда. В этом кроется одна из причин (если не главная причина), почему только обезьяна могла дать начало эволюции в сторону человека.
Важно также отметить, что в результате особого развития гаптики у обезьян «человеческая кисть, – как пишет известный советский антрополог М.Ф.Нестурх, – в общем сохранила основной тип строения от ископаемых антропоидов... Для осуществления трудовых действий, тончайших манипуляций и мастерских движений достаточным оказалось сравнительно небольшое морфологическое преобразование кисти».*
* Нестурх М.Ф. Приматология и антропогенез. М., 1960. С. 65.
Указанные глубокие прогрессивные преобразования в двигательной, особенно гаптической, сфере, подтверждаемые также данными эволюционной морфологии и палеоприматологии, сопряжены с не менее глубокими коррелятивными изменениями всего поведения. Прежде всего это относится к сенсорным функциям, в частности кожно-мышечной чувствительности руки, которая у приматов, как уже отмечалось, приобретает ведущее значение. Исключительно важным моментом является здесь взаимодействие тактильно-кинестетической чувствительности со зрением, взаимообусловленность этих сенсорных систем. Еще И.М.Сеченов подчеркивал огромное значение этого взаимодействия как фактора формирования психической деятельности человека, у которого, по словам Сеченова, «первоначальное обучение и воспитание чувств» осуществляются мышечным чувством и осязательной чувствительностью руки при сочетании движений рук со зрением. Процесс этот взаимосвязанный: по мере того как зрение «обучается» двигательной чувствительностью руки, сами движения рук все больше контролируются, корректируются и управляются зрением. По данным Фабри, и в этом отношении обезьяны составляют исключение среди млекопитающих: только у них существуют такие взаимоотношения, которые также являются одной из важнейших предпосылок антропогенеза. Ведь невозможно представить себе зарождение даже простейших трудовых операций без такого взаимодействия, без зрительного контроля за действиями рук.
Здесь важно именно то обстоятельство, что тактильно-кинестетически-оптическая чувствительность представляет у обезьян единую комплексную сенсорную систему (возникшую также на основе отсутствия антагонизма между функциями грудной конечности). Сами по себе компоненты этой системы достаточно хорошо развиты и у других позвоночных. Так, тактильно-кинестетическая чувствительность хорошо развита, например, у енота, что отчетливо проявляется хотя бы при ранее описанном открывании им запирающих механизмов. В природных условиях еноты на ощупь перебирают в воде ил и водные растения в поиске пищи, равно как и на суше, и часто производит ощупывающие движения. Однако обезьяны намного превосходят их по многообразию движений пальцев, которыми производят уже упомянутый практический анализ пищевых объектов при их очищении, расчленении и т.д. Войтонис видел именно в этой особенности питания обезьян (употребление в пищу весьма разнообразных по своим физическим качествам растительных объектов) одну из первопричин развития их ориентировочно-исследовательской деятельности, их «любопытства».
Что касается зрения, то, как уже говорилось, оно превосходно развито и у птиц. Но известное изречение Энгельса о том, что «орел видит значительно дальше, чем человек, но человеческий глаз замечает в вещах значительно больше, чем глаз орла»*, с предельной меткостью характеризует суть двух в корне различных путей развития зрения. И только зрение обезьян, развивающееся и действующее в сочетании с чувствительностью руки, особенно ее чрезвычайно подвижных пальцев, стало способным к настолько полноценному восприятию физических свойств предметов, что это оказалось достаточным и вместе с тем необходимым для выполнения трудовых операций первыми людьми. Вот почему только зрение обезьяны могло превратиться в человеческое зрение.
*Энгельс Ф. Диалектика природы. С. 135.
Предметная деятельность обезьян
Свое конкретное воплощение взаимодействие зрения и тактильно-кинестетической чувствительности рук находит в чрезвычайно интенсивной и многообразной манипуляционной активности обезьян.
Исследования, проведенные рядом советских зоопсихологов (Ладыгина-Котс, Войтонис, Левыкина, Фабри, Новоселова и др.), показали, что как низшие, так и высшие (человекообразные) обезьяны осуществляют в ходе манипулирования практический анализ объекта (расчленение, разбор его, выделение и обследование отдельных деталей и т.п.). Однако у человекообразных обезьян значительно сильнее, чем у низших, выражены синтетические («конструктивные») действия, т.е. воссоздание из частей, целого путем сближения, соединения, наслаивания предметов, их скручивания, завертывания, переплетения и т.д. Конструктивные действия встречаются в природных условиях у шимпанзе при гнездостроении.
Кроме конструктивной деятельности Ладыгина-Котс выделила еще следующие формы деятельности, выявляющиеся в ходе манипулирования предметами: ориентировочно-обследующую (ознакомительную), обрабатывающую, двигательно-игровую, сохранение предмета, отвергание его и орудийную деятельность. Количественный анализ общей структуры деятельности шимпанзе в процессе манипулирования привел Ладыгину-Котс к выводу, что по отношению к непищевым объектам чаще всего выступает ознакомительная (поверхностное ознакомление с предметами, не оставляющее на них заметных следов), обрабатывающая (углубление воздействия на предмет – царапанье, грызение, расчленение и т.п.) и конструктивная деятельность. В наименьшей степени проявлялась орудийная деятельность.
Ладыгина-Котс объясняет эти различия в удельном весе отдельных форм предметной деятельности шимпанзе особенностями образа жизни этого антропоида. Большое место, которое занимает ориентировочно-обследующая и обрабатывающая деятельность в поведении шимпанзе, объясняется разнообразием растительных кормов и сложными условиями, в которых приходится отличать съедобное от несъедобного, а также нередко сложной структурой пищевых объектов (необходимостью их расчленения, выделения съедобных или особенно вкусных их частей и т.п.).
Ладыгина-Котс обратила внимание на то, что в противоположность гнездостроительной внегнездовая конструктивная деятельность встречается редко и развита весьма слабо. Проявляется она в условиях неволи в запутывании, обматывании или переплетений, например, прутиков или веревок или в скатывании шариков из глины. При этом показательно, что такое манипулирование не направлено на получение определенного результата деятельности, а, наоборот, чаще всего переходит в деконструирование, т.е. разрушение результата деятельности (расплетение, раскручивание, расчленение и т.д.).
Минимальное развитие у шимпанзе орудийной деятельности, т.е. употребление предмета как вспомогательного средства для достижения какой-либо биологически значимой цели, Ладыгина-Котс объясняет тем, что и в естественных условиях такая форма обращения с предметами встречается крайне редко.
Действительно, несмотря на интенсивное изучение поведения человекообразных обезьян в естественных условиях, осуществленное рядом исследователей за последние годы, известны лишь отдельные случаи орудийных действий. К таким наблюдениям относятся описанные Лавик-Гудолл случаи извлечения термитов из их построек с помощью веточек или соломинок или собирания влаги из углублений в стволе дерева с помощью жеваного комка листьев. В действиях с веточками наибольший интерес представляет то обстоятельство, что прежде чем пользоваться ими как орудиями, шимпанзе (как в описанных ранее опытах Ладыгиной-Котс) отламывают мешающие листья и боковые побеги.
Даже эти, хотя лишь изредка наблюдаемые, но, безусловно, замечательные, орудийные действия дикоживущих шимпанзе являются, однако, несравненно более простыми, чем орудийные действия, искусственно формируемые у человекообразных обезьян в специальных условиях лабораторного эксперимента. Это означает, что получаемые в экспериментальных условиях данные свидетельствуют лишь о потенциальных психических способностях этих животных, но не о характере их естественного поведения. Ладыгина-Котс расценивала, самостоятельное применение орудия скорее как индивидуальную, чем видовую черту в поведении высших обезьян. Правда, как показывают полевые наблюдения, такое индивидуальное поведение может при соответствующих условиях проявляться в более или менее одинаковой мере среди многих или даже всех членов одной популяции обезьян.
Во всяком случае следует постоянно иметь в виду биологическую ограниченность орудийных действий антропоидов и тот факт, что мы имеем здесь дело явно с рудиментами прежних способностей, с угасшим реликтовым явлением, которое может, однако, как бы вспыхнуть в искусственно создаваемых условиях зоопсихологического эксперимента.
Предтрудовая предметная деятельность ископаемых обезьян
Не переоценивая орудийную деятельность современных антропоидов, нельзя одновременно не усмотреть в ней свидетельство одной из важных биологических предпосылок антропогенеза.
Надо думать, что у ископаемых антропоидов – предков человека – употребление орудий было значительно лучше развито, чем у современных человекообразных обезьян. С этой существенной поправкой можно по предметной деятельности последних, а также низших обезьян в известной мере судить о развитии предтрудовой деятельности наших животных предков и о тех условиях, в которых зародились первые трудовые действия, выполняемые с помощью орудий труда.
В этой связи важно вспомнить слова Энгельса о том, что «труд начинается с изготовления орудий»* (курсив мой. – К.Ф.). Предпосылкой этому служили в свое время, очевидно, действия, исполняемые антропоидами, подобные наблюдаемым и у современных их представителей (удаление боковых побегов веточек, отщепление лучины от дощечки и т.п.). Однако изготовляемые таким образом обезьянами (как и другими животными) орудия являются не орудиями труда, а лишь средствами биологической адаптации к определенным ситуациям.
* Энгельс Ф. Диалектика природы. С. 137.
Уже «стремление манипулировать любым предметом, не имеющим даже отдаленного сходства с пищей, способность замечать детали и расчленять сложное, – все это, – пишет Войтонис, – является первой предпосылкой проявления умения пользоваться вещью как орудием в самом примитивном смысле этого слова».* Но если мы даже допустим, что ископаемые антропоиды обладали весьма развитой способностью к употреблению орудий, то все же остается неясным, почему эта биологическая способность могла и должна была «перерасти» в качественно иную деятельность – трудовую, а тем самым почему с необходимостью на земле появился человек.
* Войтонис Н.Ю. Предыстория интеллекта. М.; Л., 1949. С. 46.
Фабри пришел на основе своих исследований к выводу, что действительно в обычных своих формах предметная, в том числе орудийная, деятельность никогда не могла бы выйти за рамки биологических закономерностей и непосредственно «перерасти» в трудовую деятельность. Очевидно, даже высшие проявления манипуляционной (орудийной) деятельности у ископаемых человекообразных обезьян навсегда остались бы не более как формами биологической адаптации, если бы у непосредственных предков человека не наступили бы коренные изменения в поведении, аналоги которых Фабри обнаружил у современных обезьян при известных экстремальных условиях. Речь идет о явлении, которое он обозначил как «компенсаторное манипулирование». Суть его заключается в том, что в резко обедненной по сравнению с естественной среде (например, в пустой клетке) у обезьян происходит коренная перестройка манипуляционной активности. В естественных условиях (или близких к ним условиях вольерного содержания) обезьяну окружает обилие пригодных для манипулирования предметов, которые распыляют внимание животных и стимулируют их к быстрой перемене деятельности.
В условиях же клеточного содержания, когда почти полностью отсутствуют предметы для манипулирования, нормальная многообразная и «рассеянная» манипуляционная деятельность обезьян концентрируется на тех весьма немногих предметах, которыми они могут располагать (или которые им дает экспериментатор). В итоге взамен разнообразных рассеянных манипуляций со многими предметами в природе животные производят не менее разнообразные, но интенсивные, сосредоточенные, длительные манипуляции с одним или немногими предметами. При этом разрозненные двигательные элементы концентрируются, что приводит к образованию значительно более сложных манипуляционных движений.
Таким образом, естественная потребность обезьян в манипулировании многочисленными разнообразными предметами компенсируется в резко обедненной предметными компонентами среде качественно новой формой манипулирования – именно «компенсаторным манипулированием».
Не вдаваясь в подробности этого сложного процесса, необходимо, однако, отметить, что только подобные новые, в корне измененные, концентрированные и углубленные действия с предметами могли служить основой зарождения трудовой деятельности. И если обратиться к фактическим природным условиям, в которых зародилось человечество, то оказывается, что они действительно ознаменовались резким обеднением среды обитания наших животных предков. В конце миоцена, особенно же в плиоцене, началось быстрое сокращение тропических лесов, и многие их обитатели, в том числе обезьяны, оказались в полуоткрытой или даже совсем открытой безлесной местности, т.е. в среде, несравненно более однообразной и бедной объектами для манипулирования. В числе этих обезьян были и близкие к предку человека формы (рамапитек, парантроп, плезиантроп, австралопитек), а также, очевидно, и наш непосредственный верхнеплиоценовый предок.
Вынужденный переход в новую среду обитания принес обезьянам, все поведение и строение которых формировалось в течение миллионов лет в условиях лесной жизни, немалые трудности, и большинство из них вымерло. Очевидно, как отмечает Нестурх, преимущества имели те антропоиды, которые смогли выработать более совершенную прямую походку на двух ногах (прямохождение) на основе прежнего способа передвижения по деревьям – по так называемой круриации. Этот тип локомоции представляет собой хождение по толстым ветвям на задних конечностях при более или менее выпрямленном положении туловища. Передние конечности лишь поддерживают при этом верхнюю часть тела.
Круриация, по Нестурху, лучше всего подготовила обезьян, сошедших на землю, к передвижению в более или менее выпрямленном положении без опоры на передние конечности, что оказалось биологически выгодным, так как эти конечности могли в результате больше и лучше использоваться для орудийной деятельности.
Из антропоидов, перешедших к жизни на открытых пространствах, уцелел, очевидно, один-единственный вид, который и стал непосредственным предком человека. Среди антропологов господствует мнение, что этот антропоид выжил, невзирая на резкое ухудшение условий жизни в начале плейстоцена, благодаря успешному использованию природных предметов в качестве орудий, а затем употреблению искусственных орудий.
Однако, как считает Фабри, такую спасательную роль, к тому же и преобразившую все поведение нашего предка и приведшую его к трудовой деятельности, орудийная деятельность смогла выполнить лишь после того, как она сама претерпела глубокую качественную перестройку. Необходимость такой перестройки была обусловлена тем, что развивавшаяся в условиях тропического леса – с его обилием разнообразных предметов – манипуляционная активность (жизненно необходимая для нормального развития и функционирования двигательного аппарата) должна была в условиях резко обедненной среды открытых пространств компенсироваться. Очевидно, тогда и возникли такие формы «компенсаторного манипулирования», которые привели к исключительно сильной концентрации элементов психомоторной сферы, что подняло орудийную деятельность нашего животного предка на качественно новую ступень.
Таким образом, высокоразвитая способность к компенсаторной перестройке предметной деятельности обеспечила выживание этого нашего предка и явилась необходимой основой для зарождения трудовой деятельности, а тем самым и появления на земле человека.
У обезьян же, оставшихся жить в лесах, естественно, не развивались компенсаторные движения, и для них были вполне достаточны прежние формы биологической адаптации, в том числе и в сфере предметной деятельности. Поэтому их орудийная деятельность осталась лишь одной из таких чисто биологических форм приспособления и не могла превращаться в трудовую деятельность. Вот почему употребление орудий у обезьян не прогрессировало, а лишь сохранялось у некоторых современных видов.
Орудия животных и орудия труда человека
Не вдаваясь в ход развития самой трудовой деятельности, отметим лишь еще несколько существенных моментов в дополнение к тому, что уже говорилось об орудийной деятельности обезьян.
Прежде всего, важно подчеркнуть, что орудием, как мы видели, может быть любой предмет, применяемый животным для решения определенной задачи в конкретной ситуации. Орудие труда же непременно должно специально изготавливаться для определенных трудовых операций и предполагает знание о будущем его применении. Они изготовляются впрок еще до того, как возникнет возможность или необходимость их применения. Сама по себе такая деятельность биологически бессмысленна и даже вредна (трата времени и энергии «впустую») и может оправдаться лишь предвидением возникновения таких ситуаций, в которых без орудий труда не обойтись.
Это значит, что изготовление орудий труда предполагает предвидение возможных причинно-следственных отношений в будущем, а вместе с тем, как показала Ладыгина-Котс, шимпанзе неспособен постичь такие отношения даже при подготовке орудия к непосредственному его применению в ходе решения задачи.
С этим связано и то важное обстоятельство, что при орудийных действиях обезьян за орудием совершенно не закрепляется его «рабочее» значение. Вне конкретной ситуации решения задачи, например до и после эксперимента, предмет, служивший орудием, теряет для обезьяны всякое функциональное значение, и она относится к нему точно так же, как и к любому другому «бесполезному» предмету. Произведенная обезьяной с помощью орудия операция не фиксируется за ним, и вне его непосредственного применения обезьяна относится к нему безразлично, а потому и не хранит его постоянно в качестве орудия. В противоположность этому не только человек хранит изготовленные им орудия, но и в самих орудиях хранятся осуществляемые человеком способы воздействия на объекты природы.
Более того, даже при индивидуальном изготовлении орудия имеет место изготовление общественного предмета, ибо этот предмет имеет особый способ употребления, который общественно выработан в процессе коллективного труда и который закреплен за ним. Каждое орудие человека является материальным воплощением определенной общественно выработанной трудовой операции.
Таким образом, с возникновением труда связано коренное изменение всего поведения: из общей деятельности, направленной на непосредственное удовлетворение потребности, выделяется специальное действие, не направляемое непосредственным биологическим мотивом и получающее свой смысл лишь при дальнейшем использовании его результатов. В этом заключается одно из важнейших изменений общей структуры поведения, знаменующих переход от естественной истории мира животных к общественной истории человечества. По мере дальнейшего развития общественных отношений и форм производства такие действия, не направляемые непосредственно биологическими мотивами, занимают в деятельности человека все большее и большее место и наконец приобретают решающее значение для всего его поведения.
Подлинное изготовление орудий труда предполагает воздействие на предмет не непосредственно эффекторными органами (зубами, руками), а другим предметом, т.е. обработка изготовляемого орудия труда должна производиться другим орудием (например, камнем). Находки именно таких продуктов деятельности (отщепи, зубила) служат для антропологов истинными свидетельствами наличия у наших предков трудовой деятельности.
Вместе с тем, по данным Фабри, при манипулировании биологически «нейтральными» предметами (а только такие могли стать орудиями труда) обезьяны хотя и воздействуют подчас одним предметом на другой (рис. 24), однако обращают при этом внимание на изменения, происходящие с объектом непосредственного воздействия, т.е. с «орудием», но не на изменения, происходящие с «обрабатываемым» («вторым») объектом, который служит не больше чем субстратом, «фоном». В этом отношении обезьяны ничем не отличаются от других животных. Напрашивается вывод, что эти предметные действия обезьян по своей сущности прямо противоположны орудийной трудовой деятельности человека, при которой, естественно, важны не столь сопровождающие ее изменения самого орудия труда, сколько изменения предмета труда (гомолога «второго объекта»). Очевидно, только в определенных экспериментальных условиях возможно переключение внимания обезьян на «второй объект».
Однако изготовление орудия труда (например, обтесывание одного камня с помощью другого) требует формирования таких специфических приемов воздействия на «второй объект», таких операций, которые привели бы к совершенно особым изменениям этого объекта, благодаря которым только он и превратится в орудие труда. Наглядный пример тому – изготовление древнейшего орудия труда первобытного человека (каменного ручного рубила, рис. 50), где усилия должны были направляться на создание заостренного конца, т.е. собственно рабочей части орудия, и широкого, закругленного верха (нуклеуса, ядрища), приспособленного к прочному удерживанию орудия в руке. Именно на таких операциях выросло человеческое сознание.
Совершенно естественно, что от создания первых орудий труда типа ручного рубила шелльской эпохи, а тем более примитивного орудия (отщепов) синантропа из дошелльской эпохи пролегал еще длинный путь до выделки разнообразных совершенных орудий труда человека современного типа (неоантропа) (рис. 51). Даже на начальном этапе развития материальной культуры неоантропа, например, кроманьонского человека, отмечается огромное разнообразие типов орудий, в том числе впервые появляются составные орудия: наконечники дротиков, кремневые вкладыши, а также иглы, копьеметалки и др. Особенно обращает на себя внимание обилие инструментов для выделки орудий. Позже появляются и такие каменные орудия, как топор или мотыга.
|
|

|
|

Рис.50. Кремневое ручное Рис. 51. Орудия позднего палеолита
рубило шелльской эпохи
Материальная культура и биологические закономерности
Знаменательно, что наряду с мощным прогрессом в развитии материальной культуры, а соответственно и психической деятельности, с начала эпохи позднего палеолита резко затормозилось биологическое развитие человека: физический тип человека приобретает очень большую устойчивость своих видовых признаков. Но у древнейших и у древних людей соотношение было обратным: при чрезвычайно интенсивной биологической эволюции, выражавшейся в большой изменчивости морфологических признаков, техника выделывания орудий труда развивалась чрезвычайно медленно.
Исходя из этого известный советский антрополог Я.Я. Рогинский выдвинул теорию «двух поворотных пунктов» в человеческой эволюции (применяется также формулировка «единый скачок с двумя поворотами»). По этой теории новые, социально-исторические закономерности появлялись у древнейших людей вместе с зарождением трудовой деятельности (первый поворот). Однако наравне с ними на протяжении большого периода продолжали действовать унаследованные от животного предка биологические закономерности. Постепенное накопление нового качества привело на завершающем этапе этого развития к крутому (второму) повороту, который состоял в том, что эти новые, социальные закономерности стали играть определяющую роль в жизни и дальнейшем развитии людей. Этот поворот в истории человечества и ознаменовался появлением человека современного типа – неоантропа. Рогинский говорит по этому поводу о снятии видообразующей роли естественного отбора и победе социальных закономерностей.
Итак, с появлением в позднем палеолите неоантропа биологические закономерности окончательно теряют свое ведущее значение и уступают свое место общественным. Рогинский подчеркивает, что только с появлением неоантропа социальные закономерности приобретают действительно господствующее значение в жизни человеческих коллективов.
Этой концепции соответствует представление о том, что первые трудовые действия должны были выполняться еще в старой (животной) форме, представленной, по Фабри, сочетанием «компенсаторного манипулирования» с обогащенной им же орудийной деятельностью. Лишь впоследствии новое содержание предметной деятельности (труд) приобрело и новую форму в виде специфически человеческих трудовых движений, не свойственных животным. Таким образом, на первых порах большому влиянию биологических закономерностей, унаследованных от животных предков человека, соответствовала внешне несложная и однообразная предметная деятельность первых людей. А это как бы маскировало свершение величайшего события – возникновения труда и вместе с ним самого человека.
Проблема зарождения общественных отношений и членораздельной речи
Групповое поведение обезьян и зарождение общественных отношений
Общественные отношения зародились в недрах первых форм трудовой деятельности. Труд с самого начала был коллективным, общественным. Это вытекало уже из того, что люди с момента своего появления на земле всегда жили коллективами, а обезьяны – предки человека – более или менее крупными стадами (или семьями). Таким образом, биологические предпосылки общественной жизни человека следует искать в стадности ископаемых высших приматов, точнее, в их предметной деятельности, выполняемой в условиях стадной жизни.
С другой стороны, труд определял с самого начала качественное своеобразие объединений первых людей. Это качественное отличие коренится в том, что даже наиболее сложная орудийная деятельность животных никогда не имеет характера общественного процесса и не определяет собой отношений между членами сообщества, что даже у животных с наиболее развитой психикой структура сообщества никогда не формируется на основе орудийной деятельности, не зависит от нее, а тем более не опосредуется ею.
Обо всем этом необходимо помнить при выявлении биологических предпосылок зарождения человеческого общества. Глубоко ошибочными являются предпринимаемые нередко попытки непосредственно вывести закономерности общественной жизни людей из закономерностей группового поведения животных. Человеческое общество не просто продолжение или усложнение сообщества наших животных предков, и социальные закономерности не сводимы к этологическим закономерностям жизни обезьяньего стада. Общественные отношения людей возникли, наоборот, в результате ломки этих закономерностей, в результате коренного изменения самой сущности стадной жизни зарождающейся трудовой деятельностью.
В поисках биологических предпосылок общественной жизни Войтонис обратился к стадной жизни низших обезьян с целью выявить те условия, в которых «появившееся у отдельных особей индивидуальное пользование орудием могло стать общественным, могло повлиять на перестройку и развитие взаимоотношений, могло найти в этих взаимоотношениях мощный фактор, стимулировавший само применение орудия».* Войтонис и Тих провели в этом направлении многочисленные исследования по выявлению особенностей структуры стада и стадного поведения у обезьян.
* Войтонис Н.Ю. Предыстория интеллекта. С. 192.
Тих придает особое значение возникновению у обезьян новой, самостоятельной и весьма мощной потребности в общении с себе подобными. Эта новая потребность, по Тих, зародилась еще на низшем уровне эволюции приматов и достигла своего расцвета у ныне живущих павианов, а также у живущих семьями человекообразных обезьян. У животных предков человека прогрессивное развитие стадности также проявилось в формировании прочных внутристадных отношений, которые оказались, в частности, особенно полезными при совместной охоте с помощью естественных орудий. Тих считает, что именно эта деятельность привела к необходимости обработки орудий охоты, а затем и к выделке примитивных каменных орудий для изготовления разнообразных охотничьих орудий.
Большое значение придает Тих и тому обстоятельству, что у непосредственных предков человека подростки должны были, очевидно, усваивать традиции и умения, сформировавшиеся у предшествующих поколений, перенимать опыт старших членов сообщества, а последние, особенно самцы, должны были проявить не только взаимную терпимость, но и умение сотрудничать, согласовывать свои действия. Всего этого требовала сложность совместной охоты с применением различных предметов (камней, палок) в качестве орудий охоты. Одновременно на этом этапе впервые в эволюции приматов возникли условия, когда появилась необходимость в обозначении предметов и без этого нельзя было обеспечить согласованность действий членов стада при совместной охоте.
Демонстрационное моделирование
Большой интерес представляет для понимания зарождения человеческих форм общения описанное Фабри «демонстрационное манипулирование» у обезьян.
У ряда млекопитающих описаны случаи, когда одни животные наблюдают за манипуляционными действиями других животных. Так, медведи нередко наблюдают за индивидуальными манипуляционными играми своих сородичей, а иногда и других животных, например выдр и бобров. Однако наиболее типично это для обезьян, которые не только пассивно наблюдают за манипуляциями другой особи, но и очень оживленно реагируют на них. Нередко случается, что одна обезьяна «провокационно» манипулирует на виду у других. Помимо демонстрационного показа объекта манипулирования и производимых с ним действий такая обезьяна часто «дразнит» другую и тем, что придвигает предмет к ней, но немедленно отдергивает его назад и с шумом «нападает» на нее, как только та протягивает к нему руку. Как правило, это повторяется много раз подряд. Такое «поддразнивание» предметом служит нередко приглашением к совместной игре и соответствует аналогичному «провокационному» поведению псовых и других млекопитающих в «трофейных» играх (см. ч. II, гл. 4), когда «заигрывание» осуществляется «вызывающим» показом игрового объекта.
В других же случаях «нарочитый» показ объекта манипулирования приводит у обезьян к несколько иной ситуации: одна особь подчеркнуто манипулирует предметом на виду у внимательно наблюдающих за ее действиями членов стада, а агрессивные проявления со стороны «актера», встречающиеся при обычном «поддразнивании», подавляются «зрителями» путем особых «примирительных» движений и поз. «Актер» же проявляет признаки «импонирования», свойственные истинному демонстрационному поведению. Такое «демонстрационное манипулирование» встречается преимущественно у взрослых обезьян, но не у детенышей.
Результатом демонстрационного манипулирования могут быть подражательные действия «зрителей», но не обязательно. Это зависит от того, насколько действия «актера» стимулировали остальных обезьян. Однако объект манипулирования всегда выступает как некий посредник в общении между «актером» и «зрителями».
При демонстрационном манипулировании «зрители» могут ознакомиться со свойствами и структурой предмета, которым манипулирует «актер», даже не прикасаясь к объекту. Такое ознакомление совершается опосредованно: происходит усвоение чужого опыта на расстоянии путем «созерцания» чужих действий.
Очевидно, демонстрационное манипулирование имеет прямое отношение к формированию «традиций» у обезьян, обстоятельно описанное рядом японских исследователей. Подобные традиции образуются в пределах замкнутой популяции и охватывают всех ее членов. Так, например, в популяции японских макаков, живших на небольшом острове, было обнаружено постепенное, но затем всеобщее изменение пищевого поведения, что выражалось в освоении новых видов пищи и изобретении новых форм ее предварительной обработки. По опубликованным данным напрашивается вывод, что это происходило на основе опосредованных игр детенышей, а затем демонстрационного манипулирования и подражательных действий обезьян.
Демонстрационное манипулирование обнаруживает все признаки демонстрационного поведения (см. ч. I, гл. 2), но при этом играет и существенную познавательную роль. Таким образом, в демонстрационном манипулировании сочетаются коммуникативные и познавательные аспекты активности: «зрители» получают информацию не только о манипулирующей особи («актере»), в действиях которого содержатся элементы «импонирования», но и (дистантно) о свойствах и структуре объекта манипулирования.
Демонстрационное манипулирование служило, по Фабри, в свое время, очевидно, источником становления чисто человеческих форм общения, так как последние зародились вместе с трудовой деятельностью, предшественником и биологической основой которой и являлось манипулирование предметами у обезьян. Вместе с тем именно демонстрационное манипулирование создает наилучшие условия для совместной коммуникативно-познавательной деятельности, при которой основное внимание членов сообщества обращено на предметные действия манипулирующей особи.
Язык животных и членораздельная речь
У современных обезьян средства общения, коммуникации отличаются не только своим многообразием, но и выраженной адресованностью, побуждающей функцией, направленной на изменение поведения членов стада. Тих отмечает также большую выразительность средств общения обезьян и их сходство с эмоциональными средствами коммуникации у человека. Однако в отличие от человека, как считает Тих, коммуникативные средства обезьян – как звуки, так и телодвижения – лишены семантической функции и поэтому не служат орудием мышления.
В последние годы коммуникационные возможности обезьян, прежде всего человекообразных, изучались особенно интенсивно, но не всегда адекватными методами. Можно, например, ссылаться на эксперименты американского ученого Д. Премака, пытавшегося с помощью системы оптических сигналов научить шимпанзе человеческому языку. По этой системе у обезьяны вырабатывались ассоциации между отдельными предметами (кусками пластика) и пищей, причем применялась методика «выбора на образец», введенная в практику зоопсихологических исследований еще в 10-х годах нашего века Ладыгиной-Котс: чтобы получить лакомство, обезьяна должна выбрать среди разных предметов (в данном случае разнообразных кусков пластика) и дать экспериментатору тот, который ей перед этим показывался. Таким же образом вырабатывались реакции на категории объектов и формировались обобщенные зрительные образы, представления подобно тем, с которыми мы уже познакомились при рассмотрении поведения позвоночных и даже пчел, но, понятно, у шимпанзе они были более сложными. Это были представления типа «большее» и «меньшее», «одинаковое» и «различное» и сопоставления типа «на», «сперва», «потом», «и» и др., на что животные, стоящие ниже антропоидов, вероятно, неспособны.
Эти опыты, как и аналогичные эксперименты других исследователей, безусловно, очень эффектно показывают исключительные способности человекообразных обезьян к «символическим» действиям и обобщениям, большие их возможности общения с человеком и, конечно, особенно мощное развитие их интеллекта – все это, правда, в условиях особенно интенсивных обучающих воздействий со стороны человека («развивающего обучения», по Ладыгиной-Котс).
Вместе с тем эти эксперименты, вопреки замыслам их авторов, никоим образом не доказывают наличия у антропоидов языка с такой же структурой, как у человека, хотя бы уже потому, что шимпанзе «навязали» подобие человеческого языка вместо того, чтобы установить с животным коммуникацию с помощью его собственных естественных средств общения. Ясно, что если судить по придуманному Премаком «пластиковому» языку как об эквиваленте подлинного обезьяньего языка, это неизбежно приведет к артефактам. Такой путь в самом принципе своем бесперспективен и не может привести к пониманию сущности языка животного, ибо данные эксперименты дали лишь феноменологическую картину искусственного коммуникационного поведения, внешне напоминающего оперирование языковыми структурами у человека. У обезьян была выработана только лишь (правда, весьма сложная) система общения с человеком в дополнение к тому множеству систем общения человека с животным, которые он создал еще начиная со времен одомашнивания диких животных.
Итак, несмотря на подчас поразительное умение шимпанзе пользоваться оптическими символическими средствами при общении с человеком и в частности употреблять их в качестве сигналов своих потребностей, было бы ошибкой толковать результаты подобных опытов как доказательства якобы принципиального тождества языка обезьян и языка человека или вывести из них непосредственные указания на происхождение человеческих форм коммуникации. Неправомерность таких выводов вытекает из неадекватного истолкования результатов этих экспериментов, при котором из искусственно сформированного экспериментатором поведения обезьян выводятся заключения о закономерностях их естественного коммуникационного поведения.
Что же касается языковых возможностей обезьян, то принципиальная невозможность обучения обезьян членораздельному языку была неоднократно доказана, в том числе и в последние годы, равно как была показана несостоятельность лингвистических выводов Премака и других авторов упомянутых экспериментов. Конечно, вопрос о семантической функции языка животных еще во многом не ясен, но нет сомнений в том, что ни у одного животного, включая и человекообразных обезьян, нет понятийного мышления. Как уже подчеркивалось, среди коммуникативных средств животных немало «символических» компонентов (звуков, поз, телодвижений и пр.), но нет абстрактных понятий, нет слов, членораздельной речи, нет кодов, обозначающих предметные компоненты среды, их качества или отношения между ними вне конкретной ситуации. Такой в корне отличный от животного способ общения мог появиться лишь при переходе с биологической на социальную плоскость развития. Одновременно, как указывал Энгельс, членораздельная речь и труд являлись главными факторами антропогенеза.
Нет ничего удивительного в том, что и язык животных характеризуется обобщенной условностью передаваемых сигналов. Это является основой любой системы коммуникации, а при переходе к социальной форме общения у первых людей это послужило биологической предпосылкой зарождения членораздельной речи в ходе их совместной трудовой деятельности. При этом только зарождающиеся общественно-трудовые отношения могли реализовать эту предпосылку, и есть много оснований думать, что первые элементы человеческой речи относились именно к этим отношениям, обозначали информацию о предметах, включенных в совместную трудовую деятельность.
В этом состоит принципиальное отличие от языка животных, который информирует прежде всего (хотя не исключительно) о внутреннем состоянии индивидуума. Как уже отмечалось, коммуникативная функция языка состоит в сплочении сообщества, индивидуальном опознавании, сигнализации о местонахождении (например, птенца или «хозяина» индивидуального участка), привлечении полового партнера, сигнализации об опасности, импонированни или запугивании и т.д. Все эти функции всецело остаются в рамках чисто биологических закономерностей.
Другое важное отличие языка животных от человеческой речи состоит в том, что язык животных всегда представляет собой «закрытую», генетически фиксированную систему, состоящую из определенного для каждого вида ограниченного количества сигналов, в то время как членораздельная речь человека является «открытой» системой, которая постоянно обогащается новыми элементами путем создания новых комбинаций из составляющих ее акустических компонентов. Поэтому человеку приходится в ходе его индивидуального развития выучить кодовые значения языка, научиться понимать и произносить их.
Формирование человеческого языка
Человеческий язык, как это имело место и в отношении материальной культуры, прошел долгий путь развития, и звуки, сопровождающие первые трудовые действия, еще не могли быть подлинными словами, обозначающими отдельные объекты, их качества или производимые с ними действия. Эти звуки вначале еще не существовали самостоятельно, а были вплетены в практическую деятельность. К тому же они непременно сопровождались жестами и выразительными интонациями, и понять их значение можно было, лишь зная ту конкретную наглядную ситуацию, в которой они возникли.
Такой «действенный разговор», осуществляемый руками, приводил, однако, как отмечает Тих, к конфликту между двумя функциями руки – действием с предметами и их обозначением, что влекло за собой передачу семантической функции голосовым органам. Тем самым было положено начало развитию самостоятельного звукового языка.
Однако врожденные звуки, жесты, мимика сохраняли свое значение начиная с первобытных людей до наших дней, правда, лишь в качестве дополнения к акустическим средствам. Все же длительное время связь этих компонентов продолжала оставаться настолько тесной, что один и тот же звуковой комплекс («праслово») мог обозначать, например, и предмет, на который указывала рука, и саму руку, и действие, производимое с этим предметом. Только после того как звуки языка отделились от практических действий, возникли первые подлинные слова. Эти слова, очевидно, обозначали предметы, и лишь значительно позже появились слова, обозначающие действия и качества.
В ходе отделения языка от непосредственно практической деятельности словесные значения становятся все более абстрактными, язык все больше выступает и как средство человеческого мышления, а не только как средство общения. Леонтьев пишет по этому поводу, что «непосредственная связь языка и речи с трудовой деятельностью людей есть то главнейшее и основное условие, под влиянием которого они развивались как носители "объективированного" сознательного отражения действительности. Обозначая в трудовом процессе предмет, слово выделяет и обобщает его для индивидуального сознания именно в этом, объективно-общественном его отношении, т.е. как общественный предмет».* То, что мышление, речь и общественно-трудовая деятельность составляют в своем зарождении и развитии единый комплекс, что при этом мышление человека могло развиваться лишь в единстве с общественным сознанием, и составляет основное качественное отличие человеческого мышления от мышления у животных. Деятельность животных и в высших ее формах всецело подчиняется естественным связям и отношениям между предметными компонентами окружающей среды. Деятельность же человека, выросшая из деятельности животных, претерпела коренные качественные изменения и подчиняется уже не столько природным, сколько общественным связям и отношениям. Это общественно-трудовое содержание и отражают слова, понятия человеческой речи.
* Леонтьев А.Н. Проблемы развития психики. 3-е изд. М., 1972. С.219.
* * *
Психика даже высших животных способна отражать лишь пространственно-временные связи и отношения между предметными компонентами среды, но не глубокие причинно-следственные связи. Психика же человека прямо или косвенно отражает также и общественные связи и отношения, деятельность других людей, равно как ее результаты, и это позволило человеку постигнуть даже недоступные наблюдению причинно-следственные связи. На этой основе стало возможным отражение в мозгу человека предметной действительности вне непосредственного отношения к ней субъекта, т.е. в сознании человека образ действительности уже не сливается с переживанием субъекта, а отражаются объективные, устойчивые свойства этой действительности.
Определяя человеческое мышление как общественно обусловленную познавательную деятельность, С.Л. Рубинштейн подчеркивал, что «мышление, в собственном смысле слова, без языка невозможно. Абстрактное мышление – это языковое, словесное мышление... Человеческое познание есть историческая категория. Оно не сводимо к моментальному акту, в котором знание возникает, чтобы тут же угаснуть. Познание в собственном смысле слова предполагает преемственность приобретаемых познаний и, значит, возможность их фиксации, осуществляемой посредством слова».* Животные лишены возможности словесного общения, а тем более словесной фиксации приобретаемых познаний и их передачи потомству с помощью языка. Этим определяется предел мышления животных, равно как их коммуникативных возможностей, и одновременно характеризуется биологическая, чисто приспособительная роль их общения. Ведь для осуществления этой роли не нужны слова, благодаря которым «впервые появляются абстрагируемые от вещей идеальные объекты мышления как "теоретической" деятельности и вместе с ними и эта последняя».**
* Рубинштейн С.Л. Бытие и сознание. М., 1957. С. 162.
** Там же. С. 162–163.
Итак, мы подошли к тому рубежу, когда на фоне филогенетической общности происхождения четко вырисовывается коренное различие между интеллектом животных и сознанием человека, а тем самым и грань между животным и человеком вообще. Переход через эту грань стал возможен и необходим в результате активного, в корне иного воздействия на природу при осуществлении трудовой деятельности. Осуществляемая с помощью орудий труда, эта деятельность опосредовала отношения ее исполнителя к природе, а это послужило важнейшей предпосылкой для преобразования досознательной психики в сознание.
Элементы опосредованного отношения к природе, к ее предметным компонентам встречаются уже в манипуляционной активности обезьян, особенно при компенсаторном манипулировании и в орудийных действиях, а также при демонстрационном манипулировании. Однако, как уже отмечалось, при сложном манипулировании, когда обезьяна воздействует одним объектом на другой, ее внимание направлено лишь на изменения, происходящие в аналоге орудия («первом объекте»), но не в объекте воздействия («втором объекте»). Подлинные же орудийные действия ситуационно обусловлены, и соответственно их познавательная ценность предельно ограничена конкретным, чисто приспособительным значением этих действий, как об этом свидетельствуют и приведенные раньше примеры. Свое всемерное развитие и полноценное познавательное значение опосредованные действия получают только тогда, когда после слияния компенсаторного манипулирования с орудийными действиями внимание переключается на обрабатываемый («второй») объект, что с необходимостью и происходит при трудовой деятельности. Именно такое и только такое опосредованное отношение к природе позволило человеку вскрыть недоступные непосредственному наблюдению существенные, внутренние взаимозависимости и закономерности природы.
Не менее существенным для преобразования досознательной психики в сознание являлось то, что опосредованное отношение к природе формировалось в ходе установления трудовых связей между работающими, через их общение друг с другом при выполнении трудовых операций. Именно таким образом возникала одновременно с сознанием в процессе трудовой деятельности членораздельная речь, послужившая средством обусловливания индивидуального сознания общественным.
Как указывал К. Маркс, сознание людей было первоначально непосредственно вплетено в их материальную деятельность и материальное общение.
О коренном отличии исторического развития человечества от биологической эволюции животных, культурного типа развития поведения от биологического Л.С.Выготский писал, что «развитие высших психических функций происходит без изменения биологического типа человека, в то время как изменения биологического типа являются основой эволюционного типа развития. Как известно и как неоднократно указывалось, эта черта составляет и общее отличие исторического развития человека». У человека, продолжает Выготский, «на первый план выступает развитие его искусственных органов – орудий, а не изменение органов и строения тела».*
* Выготский Л.С. Развитие высших психических функций. М., 1960. С. 40-41.
Но именно биологическая эволюция животных, весь ход филогенеза позвоночных, а еще ниже – беспозвоночных создали биологическую основу и предпосылки для этого небывалого в истории органического мира перехода на совершенно новый уровень развития.
Литература
1. Анохин П.К. Узловые вопросы теории функциональной системы. М.,1980.
2. Боровский В.М. Психическая деятельность животных. М.; Л., 1936.
3. Боровский В.М. Проблема инстинкта. Крымское гос. изд-во, 1941.
4. Вагнер В.А. Психология животных. 2-е изд. М., 1902.
5. Вагнер В.А. Биологические основания сравнительной психологии. Т. 1, 2. Спб.; М., 1910-1913.
6. Вагнер В.А. Сегментарная психология // Биопсихология. Новые идеи в биологии. Сб. 6. Спб., 1914.
7. Вагнер В.А. Возникновение и развитие психических особенностей. Этюды по сравнительной психологии. Вып. 1–9. Л., 1925-1929.
8. Войтонис НЮ. Предыстория интеллекта. М.; Л., 1949.
9. Выготский Л. С. Предисловие к русскому изданию книги Кёлера «Исследование интеллекта человекоподобных обезьян» // Выготский Л.С. Собр. соч. Т. 1. М., 1982. № 10. Дарвин Ч. Инстинкт // Соч. Т. 3. М., 1963.
11. Дарвин Ч. Выражение эмоций у человека и животных // Соч. Т. 5. М., 1953.
12. Дембовский Я. Психология животных. М., 1959.
13.Дембовский Я. Психология обезьян. М., 1963.
14.ДьюсбериД. Поведение животных. Сравнительные аспекты. М., 1981.
15. Захаров А.А. Внутривидовые отношения у муравьев. М., 1972.
16. Кёлер В. Исследование интеллекта человекоподобных обезьян. М., 1930.
17. Крушинский Л.В. Биологические основы рассудочной деятельности: эволюционные и физиологогент. аспекты поведения. М.,1986.
18.Лавик-ГудоллДж. В тени человека. М., 1974. № 19 Лавик-Гудолл Дж. Невинные убийцы. М., 1977.
20. Ладыгина-Котс Н.Н. Исследование познавательных способностей шимпанзе. М., 1923.
21. Ладыгина-Котс Н.Н. Дитя шимпанзе и дитя человека в их инстинктах, эмоциях, играх, привычках и выразительных движениях. М., 1935.
22. Ладыгина-Котс Н.Н. Элементарное мышление животных // Вопросы психологии. 1955. № 3.
23. Ладыгина-Котс Н.Н. Развитие психики в процессе эволюции организмов. М., 1959.
24. Ладыгина-Котс Н.Н. Конструктивная и орудийная деятельность высших обезьян (шимпанзе). М., 1959.
25. Ладыгина-Котс Н.Н. Предпосылки человеческого мышления. М., 1965.
26. Ламарк Ж.Б. Философия природы // Избранные произведения. Т. 1. М., 1955.
27. Леонтьев А.Н. Проблемы развития психики. 3-е изд. М., 1972.
28. Линден Ю. Обезьяны, человек и язык. М., 1981.
29. Лопатина Н.Г. Сигнальная деятельность в семье медоносной пчелы. Л., 1971.
30. Лоренц К. Кольцо царя Соломона. М., 1970.
31. Лоренц К. Человек находит друга. М., 1971.
32. Мазохин-Поршняков Г.А. Обучаемость насекомых и их способность к обобщению зрительных стимулов // Энтомологическое обозрение. Т. 47. М., 1968.
33. Мазохин-Поршняков Г.А. Только ли инстинкт управляет поведением насекомых? // Природа. 1970. № 5.
34. Мак-Фарленд Д. Поведение животных. Психобиология, этология и эволюция. М., 1988.
35. Мешкова Н.Н. Об онтогенезе игрового поведения у некоторых хищных и парнокопытных млекопитающих // Зоологический журнал. 1970. Т. 49. Вып. 6.
36. Мешкова Н.Н. Познавательная функция манипуляционной активности серой крысы // Вести. Моск. ун-та. Сер. 14, Психология. 1981. № 3.
37. Мешкова Н.Н., Шутова М.И. Особенности психической деятельности серой крысы // Новые материалы по биологии серой крысы. М., 1990.
38. Нестурх М.Ф. Происхождение человека. М., 1958.
39. Нестурх М.Ф. Приматология и антропогенез. М., 1960.
40. Новоселова С.Л. Исследование в области экспериментального мышления шимпанзе // Вопросы экспериментальной психиатрии. М., 1969.
41. Орбели Л.А. Вопросы высшей нервной деятельности. М.; Л., 1949.
42. Панов Е.Н. Знаки, символы, языки. 1983.
43. Прайор К. Несущие ветер. М., 1981.
44. Понугаева А.Г. Импринтинг(запечатлевание).Л., 1973.
45. Промптов А.Н. Очерки по проблеме биологической адаптации поведения воробьиных птиц. М.; Л., 1956.
46. Протопопов В. П. Исследование высшей нервной деятельности в естественном эксперименте. Киев, 1950.
47. Рогинский Я.Я. Проблемы антропогенеза. М., 1969.
48. Рубинштейн С.Л. Основы общей психологии: В 2 т. М., 1989.
49. Рулье К.Ф. Избранные биологические произведения. М., 1954.
50. Северцов А. Н. Эволюция и психика. М., 1922.
51. Северцов А. Н. Главные направления эволюционного процесса.
3-е изд. М., 1967.
52. Скребицкий Г.А. Отношение чайки к гнезду, яйцам и птенцам // Рефлексы, инстинкты и навыки. Психологические исследования. Т.2.М., 1936.
53. Тинберген Н. Поведение животных. М., 1969.
54. Тинберген Н. Мир серебристой чайки. М., 1975.
55.ТихН.А. Ранний онтогенез поведения приматов. Сравнительно-психологическое исследование. Л.,1966.
56. Тих Н.А. Предыстория общества. Л., 1970.
57. Тушмалова Н.А. Основные закономерности эволюции поведения беспозвоночных // Физиология поведения. Л., 1987.
58. Фабр Ж.-А. Жизнь насекомых. М., 1963.
59. Фабри К.Э. Хватательная функция руки приматов и факторы ее эволюционного развития. М., 1964.
60. Фабри К.Э. О некоторых основных вопросах этологии // Бюллетень Московского общества испытателей природы. Отдел биологический. 1967. Т. 72. Вып. 5.
61. Фабри К.Э. В.А.Вагнер и современная зоопсихология // Вопросы психологии. 1969. №6.
62. Фабри К.Э. К проблеме игры у животных // Бюллетень Московского общества испытателей природы. Отдел биологический. 1973. Т. 78. Вып. 3.
63. Фабри К.Э. О подражании у животных // Вопросы психологии. 1974. № 2.
64. Фабри К.Э. Орудийные действия животных. М., 1980.
65. Фабри К.Э. Игра у животных. М., 1985.
66. Фирсов Л.А. Память у антропоидов. Л., 1972.
67. Фирсов Л.А. Поведение антропоидов в природных условиях. Л., 1977.
68. Фосси Д. Гориллы в тумане. М., 1990.
69. Фресс П., Пиаже Ж. Экспериментальная психология. Вып. 1,2. М., 1966.
70. Фриш К. Из жизни пчел. М., 1966.
71. Хайнд Р. Поведение животных. М., 1975.
72. Шаллер Дж. Б. Год под знаком гориллы. М., 1968.
Приложение
Игры животных и игры детей (сравнительно-психологические аспекты)*
* Вопр. психологии. 1982. № 3. С. 26–34.
Игры животных уже давно служат объектами интенсивных зоопсихологических и этологических исследований, однако в изучении сравнительно-психологических аспектов игры на сегодняшний день в сущности мало что изменилось со времен известных работ Н. Н. Ладыгиной-Котс. Вместе с тем вопрос о возможностях (и необходимости) сопоставления игр детенышей животных и игр детей представляет, конечно, большой теоретический и практический интерес, в частности, для детской психологии и дошкольной педагогики.
В этом плане здесь приводятся некоторые выводы, вытекающие из разработанной нами концепции игры животных как развивающейся психической деятельности и предложенной нами классификации игр животных. Согласно этой концепции, игры животных представляют собой не особую категорию поведения, как принято считать, а основное содержание определенного периода онтогенеза – ювенильного (преадультного, игрового), свойственного только высшим позвоночным (птицам и млекопитающим). Иными словами, игра – это совокупность специфически преадуальных проявлений общего процесса развития поведения в онтогенезе животных, это – адуальное поведение в процессе его становления. По сравнению же с предшествующим, ранним постнатальным периодом игра составляет новое содержание поведения, определяющее дозревание первичных элементов поведения и их преобразование в поведение взрослых (половозрелых) животных.
При этом, как показали результаты приведенного нами 35-летнего исследования манипуляционного поведения разных видов млекопитающих, в ходе игры развиваются не целиком поведенческие акты взрослых животных, а составляющие их сенсомоторные компоненты. Эти компоненты, развитие которых началось еще в эмбриональном (пренатальном) периоде онтогенеза, подвергаются в игровом периоде глубоким функциональным изменениям, и лишь в качественно преобразованном виде они становятся основными частями «окончательного» (взрослого) поведения. Следовательно, имеет место не «просто» перерастание игровых действий во взрослые, а формирование из элементов этих действий качественно нового поведения взрослых животных. Этот процесс преобразования сенсомоторных компонентов поведения совершается на основе накопления фундаментального индивидуального опыта, чем определяется познавательная функция игры у животных и вообще значение игры в онтогенезе психики животных.
Упомянутые функциональные преобразования компонентов поведения соответствуют общим морфофункциональным преобразованиям и, так же как и те, носят характер расширения, усиления или смены функции. С точки зрения психологического анализа наибольший интерес представляют ниже рассматриваемые явления субституции (замещения объектов воздействия), поскольку этими преобразованиями устанавливаются наиболее существенные новые связи с компонентами окружающей среды, чем обусловливается качественное обогащение содержания психического отражения. Можно даже сказать, что в целом психическое содержание игровой активности, а тем самым развитие всей психической деятельности в ювенильном периоде онтогенеза определяются установлением молодым животным различных субституционных отношений и связей с компонентами среды. Их можно определить как «преадаптивно-компенсаторные» связи, замещающие, предваряющие и имитирующие жизненные ситуации и взаимоотношения взрослых животных. В этом смысле вся игровая активность представляет собой преадаптивную субституцию взрослого поведения.
Исходя из понимания игры как развивающейся психической деятельности, мы классифицировали формы игры у животных именно на основе характера устанавливаемых ими связей с компонентами среды, выделив ряд категорий игр. При их описании мы пользуемся следующими условными обозначениями: И – индивид; И(И) – индивид, замещающий естественного игрового партнера; О – биологически значимый объект; 0(0) – объект, замещающий биологически значимый объект; 0(И) – объект, замещающий игрового партнера; С – субстрат игровой активности. Стрелками обозначаются активно устанавливаемые играющими животными связи.
ОДИНОЧНЫЕ (ИНДИВИДУАЛЬНЫЕ) ИГРЫ
Неманипуляционные игры
КАТЕГОРИЯ 1. Игры типа И®С. (Локомоционные игры; устанавливаются только непосредственные связи индивида с субстратом игровой активности.)
Манипуляционные игры
КАТЕГОРИЯ 2. Игры типа И®О. (Манипуляционные игры с пищей, гнездостроительным материалом и тому подобными биологически значимыми объектами; устанавливаются непосредственные связи индивида с объектом игры.)
КАТЕГОРИЯ 3. Игры типа И®О(О). (Устанавливаются связи, аналогичные связям в категории 2.)
КАТЕГОРИЯ 4. Игры типа И«О(И). (Устанавливаются связи, аналогичные связям в категориях 2 и 3, но иногда встречаются также элементы имитации обратной связи.)
СОВМЕСТНЫЕ ИГРЫ
Неманипуляционные игры
КАТЕГОРИЯМ. Игры типа И«И. (Устанавливаются непосредственные обоюдные связи с игровыми партнерами.)
КАТЕГОРИЯ 6. Игры типа И«И(И). (С замещающим игровым партнером – животным или человеком – устанавливаются связи, аналогичные связям в категории 5.)
Манипуляционные игры
КАТЕГОРИЯ 7. Игры типа И – О – И. (Соответствуют играм категории 2, но с объектом устанавливаются связи одновременно двумя или более партнерами.)
КАТЕГОРИЯ 8. Игры типа И = О = И. (Манипуляционные игры, при которых предмет, однако, не замещает биологически значимый объект или игрового партнера, а служит средством общения между партнерами; между последними при этом устанавливаются сложные, опосредствованные предметом, обобщенные связи.)
Разумеется, в конкретном поведении молодых животных приведенные категории встречаются часто одновременно в различных сочетаниях или переходят друг в друга. Например, совместные неманипуляционные игры, будучи по форме выполнения локомоторными, тут же в пределах тех же категорий 5 или 6, становятся по форме манипуляционными (точнее, локомоционно-манипуляционными), как только игровые партнеры вступают в физический контакт друг с другом.
Конечно, существенная разница состоит при этом в том, что, в отличие от «обычного» манипулирования предметами, манипуляционные движения направляются здесь не просто на физическое тело, а на активного индивида, на игрового партнера, отвечающего на эти движения также манипуляционными действиями, в результате чего между животными возникает взаимодействие в условиях общения. А это значит, что в пределах общей для всех категорий морфофункциональной формы появилось качественно новое психическое содержание, что и делает необходимым выделение специальных категорий 5 и 6. Если же индивида (игрового партнера) рассматривать как всего-навсего компонент среды, то выделение этих категорий делался излишним, и они целиком вольются в категорию 2. Не менее четко психическое содержание выступает в компенсаторных манипуляционных играх категорий 3 и 4. Таким образом, предлагаемая схема отражает прежде всего психические аспекты, психические векторы и связи игровой активности животных.
Отмеченные выше субституционные преобразования проявляются в разных категориях по-разному и в различной степени – в наибольшей они выражены в компенсаторных играх (категории 3, 4, 6). Однако, как уже говорилось, все игры являются в своей сущности субституционными, и поэтому игра, вне сомнения, представляет собой самое лабильное, что вообще существует в поведении животных. Здесь почти все может заменяться и замещаться, вся активность животного строится на компенсации, транспозиции, опосредованности воздействий (категория 8) и других субституционных проявлениях.
На игровом этапе онтогенеза самое главное для развивающегося животного – научиться устанавливать связи, «отработать» это умение, а лишь затем «уточнить» окончательные (биологически значимые) объекты воздействия и вовлекать их в сферу своей (теперь уже направленной) двигательной активности. Играя, молодое животное должно научиться устанавливать разнообразные связи с максимальным биологическим эффектом, но с наименьшей тратой сил и времени, да притом в постоянно меняющихся внешних условиях, в условиях постоянно возникающих ситуаций новизны. В этом и заключается значение игры для молодых животных.
Из изложенного вытекает, что игрой можно назвать лишь такую форму активности, которой присущи признаки субституции. Все компоненты ювенильного поведения, лишенные таких признаков (например, движения, выполняемые при еде), являются по своей природе персистентными компонентами раннего постнатального поведения. Эти компоненты претерпевают на ювенильном этапе онтогенеза под влиянием игровой активности определенные модификационные (но не субституционные!) изменения и в результате превращаются в элементы взрослого поведения животных без существенных качественных преобразований.
В итоге вырисовывается следующая картина. Первоначально устанавливаемые детенышем связи с компонентами среды (и соответственные формы психического отражения) являются хотя и элементарными, примитивными, но при этом непосредственно биологически эффективными и жизненно необходимыми. Установление этих связей (И – С, И – О, И = И) – это поведение •«всерьез». То же самое имеет место при соответствующем установлении связей взрослым животным, только это уже зрелое, развитое поведение качественно более высокого уровня. Вклинивающаяся же между ними игровая активность молодых животных – это поведение «не всерьез», поскольку оно не дает непосредственного биологического эффекта (ввиду субституционального характера устанавливаемых связей).
Разумеется, наряду с игровыми животные устанавливают на ювенильном этапе развития также жизненно необходимые связи «всерьез», которые обеспечивают их коренные биологические потребности. Но ювенильный период тем и отличается, что наряду с такими устанавливаются непосредственно «бесполезные», связи, которые и определяют сущность этого периода. При этом самое важное состоит в том, что формирование взрослого поведения, поведения «всерьез», происходит не путем усложнения жизненно необходимых компонентов поведения молодого животного, например поедания (но не добывания!) пищи (эти компоненты только «дозревают»), а путем развертывания и совершенствования как раз биологически «бесполезных», «несерьезных», «пробных» – одним словом, субституционных игровых действий.
Взрослое поведение, следовательно, не «вырастает» прямо и непосредственно из форм раннего постнатального поведения; процесс прямолинейного развития прерывается здесь на ювенильной, игровой стадии, и после глубокой качественной перестройки разрозненные первоначальные элементы поведения в преобразованном, обогащенном, обновленном состоянии вновь группируются по прежним направлениям развития (И – О, И = И, И – О – И). В итоге взрослое поведение отличается значительно большей гибкостью, лабильностью, чем первоначальное постнатальное. Вот почему мы и называем игру развивающейся психической деятельностью.
Все это относится, разумеется, только к высшим позвоночным, и именно в этом кроется причина исключительной пластичности их поведения. У других же животных взрослое поведение действительно непосредственно «вырастает» из первичного постнатального поведения, которое формируется в раннем постэмбриональном онтогенезе на основе созревания врожденных компонентов поведения, врожденного узнавания и раннего опыта (первичного облигатного научения и элементов примитивного факультативного научения). И совсем иного рода закономерности развития поведения у животных, претерпевающих в онтогенезе метаморфозы (земноводные, насекомые и др.), когда уже личиночные формы ведут своего рода «взрослый» образ жизни (за исключением репродуктивной функции), который, однако, в корне отличается от подлинно взрослого образа жизни адультных животных (имаго). Таким образом, не приходится говорить о неких «универсальных» закономерностях развития поведения в онтогенезе животных.
К характеристике игровой активности следует добавить, что с помощью разработанной нами системы классификации элементов манипулирования удалось установить, что при переходе от раннего постнатального периода к ювенильному появление игровой активности приводит к подлинному скачку в моторной сфере: резко увеличивается число элементов манипулирования и число объектов манипулирования. Эти количественные изменения в обращении животных с предметами сопряжены с коренными качественными изменениями, находящими свое выражение в установлении принципиально новых связей с компонентами среды. Следовательно, увеличение количества элементарных двигательных компонентов поведения детеныша приводит к новому качеству психического отражения.
Важно также подчеркнуть, что весь процесс развития поведения (а в связи с ним и психического отражения) протекает у высших позвоночных в виде отрицания: первичные связи, установленные детенышем с компонентами окружающего мира в раннем постнатальном периоде, частично замещаются в ювенильном периоде более сложными, но временными и биологически непосредственно «бесполезными», субституционными игровыми связями, которые затем (у взрослых животных) также элиминируются и заменяются вновь связями исходного типа, но наполненными качественно новым содержанием. Эти индивидуальные связи, следовательно, гомологичны элементарным первичным связям, но вторично формируются уже на новом, более высоком уровне. (Повторные качественные изменения, как уже отмечалось, происходят и в пределах категорий 1, 2 и 5, хотя здесь и имеет место, казалось бы, «прямолинейное» развитие.)
Изложенное иллюстрирует рис. 52, изображающий соответствующие переходы с одного «этажа» онтогенеза на другой (цифры – номера категорий игр, остальные условные обозначения приведены выше).
Мы видим, таким образом, что у высших позвоночных онтогенез поведения и психики представляют собой в целом не плавный непрерывный процесс, а благодаря вклиниванию игрового периода процесс, прерываемый периодом отрицания первичного содержания. Подтверждается и правомерность предложенной нами периодизации онтогенеза психической деятельности животного с выделением особого, качественно отличного игрового периода, благодаря которому и возникает эта диалектическая прерывистость процесса развития психической деятельности в онтогенезе животных.
Рис. 52. Развитие форм игровой активности у млекопитающих (объяснение см. в тексте)
* * *
При сопоставлении игр животных с играми детей исследователь наталкивается на те же трудности, что и при сравнении поведения животных и человека вообще. Трудности эти возникают из-за необходимости всестороннего учета коренных, качественных отличий поведения человека от такового даже наиболее высокоорганизованных животных, например шимпанзе. Вместе с тем возможность и даже необходимость такого сравнения определяются тем, что поведение человека наряду с ведущими, социально обусловленными включает в себя и биологические, унаследованные от наших животных предков компоненты и признаки, которые являются до своей форме в такой же степени общими с таковыми высших животных, в какой у нас с ними общими являются строение и функции организма. Сюда относятся, в частности, биологические механизмы поведения (врожденные пусковые механизмы, процессы смещения активности, запечатления и др.), определяющие во многом общую с животными форму протекания ряда важных поведенческих актов.
Основную предпосылку научно достоверного сопоставления поведения человека и животных мы видим в том, что при всех без исключения сравнительно-психологических исследованиях необходимо прежде всего исходить из четкого различия формы и содержания поведения. Содержание поведения человека всегда отличается качественно от такового животных, причем эти специфически человеческие признаки его поведения возникли в результате антропогенеза вместе с зарождением трудовой деятельности, членораздельной речи и общества, в то время как поведение животных осталось всецело биологически детерминированным, никогда и ни в одном случае не выходило за пределы биологических закономерностей, чем и определяется чисто биологическое содержание этого поведения. Поэтому содержание поведения животных и человека принципиально несопоставимо, точнее сказать – здесь возможно только сравнительное выявление различий.
Другое дело – форма человеческого поведения, которая в большинстве случаев, правда, также претерпела в ходе исторического развития социально обусловленные качественные изменения и в результате приобрела специфически человеческие черты, но в отдельных случаях сохранила в большей или меньшей степени животнообразный вид. Вот здесь и открывается плодотворное поле деятельности для сравнительной психологии, для выявления генетически обусловленных признаков сходства или даже общности в поведении животных и людей. Иными словами, если не считать некоторых примитивных поведенческих актов, сравнительно-психологический поиск общих для человека и животных признаков поведения (или признаков гомологического сходства) возможен только в отношении форм поведения, а также первичных сенсомоторных компонентов биологических механизмов поведения, но не его содержания.
Сказанное всецело относится и к сравнительно-онтогенетическому анализу поведения человека и животного, поскольку содержание поведения человека не только во взрослом состоянии, но и на всех этапах его постнатального развития качественно отличается от такового животных. Однако в некоторых случаях на начальных этапах онтогенеза это качественно новое, специфически человеческое, психическое содержание еще сохраняет некоторое время старую, унаследованную от наших животных предков и поэтому во многом общую с животными форму. Это сказывается и на общем ходе развития поведения.
Очевидно, мы имеем здесь дело с частным проявлением общей закономерности – развитием нового содержание при первоначальным сохранении старой формы до ее замены новой адекватной формой. Есть основания полагать, что подобные соотношения определили и начальный этап антропогенеза, точнее – условия зарождения трудовой деятельности.
В некоторых ранних играх детей младшего возраста, которые только и можно сопоставить с играми детенышей животных, обнаруживаются определенные компоненты, гомологичные формам игровой активности детенышей высших животных, хотя содержание и этих игр уже социально детерминировано. У детей же более старшего возраста почти всецело специфически человеческими становятся и формы игры. Об этой специфике содержания детской игры, в частности, в раннем возрасте дают представление обстоятельные исследования М.Я. Басова, Л.С. Выготского, С.Л. Рубинштейна, А.Н. Леонтьева, Д.Б. Эльконина и труды других советских психологов, посвященные играм детей. Так, например, А.Н. Леонтьев усматривал «специфическое отличие игровой деятельности животных от, игры, зачаточные формы которой мы впервые наблюдаем у детей предшкольного возраста», прежде всего в том, что игры последних представляют собой предметную деятельность. Последняя, «составляя основу осознания ребенком мира человеческих предметов, определяет собой содержание игры ребенка».*
* Леонтьев А.Н. Проблемы развития психики. М., 1959. С. 384–385.
Важнейшие качественные отличия даже ранних игр детей связаны с активным сознательным воспитанием ребенка взрослыми в условиях постоянного общения с «им, его целенаправленным приобщением к искусственному миру предметов человеческого обихода и социальной ориентации его поведения. В этом же русле совершается и специфически человеческое овладение ребенком речью – процесс несопоставимый с развитием коммуникативных систем животных. Именно путем установления таких связей начинается передача от одного поколения к следующему общественно исторического опыта. При этом, как показала М.И. Лисина, общение ребенка со взрослым при их совместной игровой предметно-манипулятивной деятельности является важнейшим условием общего психического развития ребенка. Экспериментальную разработку и обстоятельный теоретический анализ разнообразных вопросов, связанных с процессом присвоения ребенком общественного опыта, осуществляли А. А. Люблинская, Н. Н. Поддъяков и другие психологи.
Не останавливаясь на конкретных играх детей и детенышей, рассмотрим лишь некоторые общие сравнительно-онтогенетические аспекты гомологического сходства форм развития поведения.
Признаки такого общего гомологического сходства проявляются прежде всего в том, что как у детенышей животных, так и у детей в ходе онтогенеза происходит глубокая перестройка устанавливаемых ими связей с компонентами окружающего мира, в корне меняется отношение к ним. И в том и в другом случае переход от доигрового к игровому периоду характеризуется трансформацией первичной двигательной активности, особенно при обращении с предметами, резко расширяется сфера предметной деятельности, качественно меняются способы манипулирования предметами и отношение к ним, причем как у животных, так и у детей эти изменения носят прежде всего субституционный характер (наряду с проявлениями расширения, усиления и смены функции).
Разумеется, структура процесса становления и развития игровой активности ребенка носит значительно более сложный характер, чем у животных. Об этом свидетельствуют, например, данные Р.Я. Лехтман-Абрамович, согласно которым доигровые предметные действия («преддействия») сначала переходят в «простые результативные повторные предметные действия», затем в «соотносимые действия», при которых ребенок выявляет пространственные соотношения. Эти игровые действия в свою очередь сменяются «функциональными действиями», которые характеризуются тем, что ребенок распространяет свою активность на новые предметы. В результате этого обобщается накопленный двигательный опыт, начинается (путем сравнения и различия) ознакомление с функциями объектов и устанавливаются некоторые простые и общие их признаки. Продолжением этого исследования служит работа Ф.И. Фрадкиной, посвященная выделению качественных различий между этапами развития уже самой игры в раннем детстве, которые в свою очередь являются предпосылкой развитой, творческой игры. Развитие структуры предметно-опосредованных действий ребенка в раннем детстве и изменение содержания опыта его деятельности были подробно изучены С.Л. Новоселовой.
Эта сложность, «многоступенчатость» хода развития, несомненно, обусловливает специфические особенности игры в дошкольном возрасте, причем в отношении не только ее наполнения социально детерминированным содержанием, но и развития особой структуры, адекватной сугубо человеческой форме игры дошкольников. В этой связи следует напомнить и о том, что А.В. Запорожец выделяет наряду с общевозрастным процессом развития процесс функционального развития, которое связано с овладением ребенком отдельными знаниями и способами действий, в то время как возрастное развитие характеризуется образованием новых психофизиологических уровней, формированием новых планов отражения действительности, изменением ведущего отношения ребенка к действительности, переходом к новым видам деятельности. Этим содержание психического развития ребенка, конечно, также в корне отличается от содержания этого процесса у детенышей животных. К тому же функциональное развитие, безусловно, протекает у первых в качественно совершенно иных формах, чем у последних.
Большой интерес представляет появление в ходе игровой активности обобщенного отношения к предметам и условность устанавливаемых при этом связей. «Игровое действие, – указывает А. Н. Леонтьев, – всегда обобщено, это есть всегда обобщенное действие». И далее: «Именно обобщенность игровых действий есть то, что позволяет игре осуществляться в неадекватных предметных условиях».* Правда, «в игре не всякий предмет может быть всем. Более того, различные игровые предметы-игрушки выполняют в зависимости от своего характера различные функции, по-разному участвуют в построении игры».** Как общую закономерность все это можно отнести к играм животных, которых также характеризует обобщенность отношения к игровым объектам (особенно в категориях 3, 4, 6 и 8). Однако эта общность форм обращения с предметами (и игровыми партнерами) кончается с появлением развитых детских игр с игрушками (в дошкольном возрасте), которые и имел в виду А.Н. Леонтьев. Эти игры уже качественно иные и по форме. К тому же у ребенка с самого начала формируется социально обусловленное обобщенное отношение к окружающему, чем определяется качественное отличие содержания даже самых ранних игр детей.
* Леонтьев А.Н. Проблемы развития психики. М., 1959. С. 395.
** Там же. С. 396.
Что же касается отмеченного А. Н. Леонтьевым дифференцированного применения различных предметов в игровых действиях, то, в отличие от детей, у животных пригодность предмета служить «игрушкой» и конкретный способ его применения зависят от того, является ли данный предмет носителем того или иного ключевого раздражителя, т.е. детерминанты инстинктивного поведения. Иными словами, у высших животных развитие и совершенствование инстинктивных начал, их обрастание благоприобретаемыми компонентами, совершаются преимущественно путем упражнения с предметами, замещающими истинные объекты инстинктивных действий. Особенно наглядно это проявляется в играх категорий 3, 4 и 8, при которых ознакомление с миром осуществляется именно путем пробных действий с такими замещающими объектами, т.е. безопасными и бесполезными для «серьезных дел». Возможность такого преадаптационного «проигрывания» аналогов ситуаций предстоящей взрослой жизни обеспечивается тем, что оно выполняется в «зоне безопасности» – под защитой родительской особи.
Важно также подчеркнуть, что, по А.Н. Леонтьеву, на начальном этапе развития ребенка игра зарождается из предметных действий неифового типа и на первых порах является подчиненным процессом, зависящим от этих действий. Затем же игра становится ведущим типом деятельности, что связано с овладением «широким миром человеческих предметов». Тем самым перед ребенком открывается «путь осознания человеческого отношения к предметам, т.е. человеческого действия с ними».* Д.Б. Эльконии также указал на то, что доигровой период развития характеризуется первичной манипуляционной активностью, элементарными манипуляционными упражнениями, на основе которых возникают собственно игровые действия ребенка, определяемые Элькониным как действия с предметами, замещающими объекты поведения взрослых. Большое значение приобретает при этом также усложнение этих действий в сторону воспроизведения жизненных связей, характерных для определенных ситуаций жизни взрослых.
* Там же. С. 385.
Если учесть отмеченные специфические особенности развития игрового поведения ребенка, то сопоставление с развитием игровой активности животных, отраженным в приведенной схеме, показывает, что существует определенное сходство в том, что и у детей и у детенышей высших животных диалектика развития поведения проявляется в субституционных преобразованиях, которые обусловливают замену устанавливаемых с компонентами среды первичных связей вторичными, «имитационными» и «суррогативными», т.е. игровыми связями, которые затем в свою очередь заменяются связями, присущими взрослому поведению.
В этих преобразованиях, в этом отрицании отрицания устанавливаемых связей, о котором уже говорилось выше, мы усматриваем общефункциональное значение игры, точнее, игрового периода, который возник на высших этапах эволюционного процесса как промежуточное звено между начальным, постнатальным и адультным этапами онтогенеза животных. Расчленение прежде непрерывного хода онтогенеза поведения промежуточным игровым периодом было на высшем уровне эволюции животного мира совершенно необходимо для дальнейшего прогрессивного развития психической деятельности высших животных, поскольку включение в онтогенез периода установления временных субституционных связей с компонентами среды обеспечило возможность существенного расширения диапазона активности и придания поведению большей лабильности, полноценного развития всех двигательных и познавательных возможностей в ходе многостороннего воздействия на компоненты окружающего мира и активного ознакомления с ним. Ясно, что такое коренное «расширение кругозора» на основе максимальной интенсификации двигательной активности являлось (и является) необходимой предпосылкой для усиленного накопления разнообразного жизненно важного индивидуального опыта.
В процессе своего становления человек унаследовал от своих животных предков это высокоэффективное, обладающее большой адаптивной ценностью достижение эволюции, и этим обусловлена гомологичность общего хода онтогенетического развития поведения у высших животных и у человека. Однако в онтогенезе человека открываются практически безграничные возможности многократного преобразования форм игровой активности, в то время как даже у наиболее развитых животных адультное поведение фиксируется в указанных нескольких конечных руслах, изображенных на схеме. Поэтому, хотя у человека поведение развивается отчасти по унаследованным от животных жестким руслам, оно в решающих сферах его деятельности не претерпевает на адультном этапе присущее животным сужение, а, наоборот, дивергентно развивается во все новых формах. В результате поведение человека развивается в конечном итоге в целом в значительно более широком диапазоне, чем на предшествующих этапах онтогенеза.
Не вдаваясь в подробности, мы коснулись здесь лишь некоторых сравнительно-онтогенетических аспектов развития поведения и психики животных и человека, опираясь при этом на нашу концепцию игры как развивающейся психической деятельности. Многие вопросы еще нуждаются в обстоятельном изучении, а приведенные здесь обобщения носят во многом предварительный характер. Разработка этой чрезвычайно сложной проблемы, без сомнения, имеет большое значение и для решения ряда насущных практических задач дошкольной педагогики.
Научное наследие А.Н. Леонтьева и вопросы эволюции психики*
* А.Н.Леонтьев и современная психология / Под ред. А.В.Запорожца и др. М., 1983. С. 101–117.
Научное познание психики человека начинается с зоопсихологии. «Ясно, что исходным материалом для разработки психических фактов должны служить, как простейшие, психические проявления у животных, а не у человека», – подчеркивал еще 110 лет тому назад И.М.Сеченов, выделив эти слова в качестве одного из основных тезисов своего труда «Кому и как разрабатывать психологию». Правомерность этого требования и сейчас ни одним серьезным ученым не ставится под сомнение, более того, в наше время эта задача злободневна, как никогда. Познание психики невозможно без познания закономерностей ее становления и развития, без выявления ее предыстории и этапов развития психического отражения, начиная от его первичных, наиболее примитивных форм до высших проявлений психической активности животных и сопоставления последних с психическими процессами у человека. Эту поистине гигантскую задачу должна и призвана решать зоопсихология.
Разумеется, при изучении этих вопросов перед исследователем встают неимоверные трудности, в частности, очень трудно получить представление о конкретных предпосылках и условиях возникновения человеческого сознания, о непосредственной предыстории зарождения трудовой деятельности, членораздельной речи, человеческого мышления и социальной жизни. Ведь зоопсихолог может опереться на поведение только современных животных. Но сравнительно-психологический анализ их поведения и поведения человека открывает вполне реальные возможности для решения обозначенных задач и выявления качественных особенностей и отличий человеческой психики. Необходимо только учесть, что история человечества – миг по сравнению с полутора миллиардами лет развития животной жизни на земле, что в результате этого грандиозного процесса эволюции возникло огромное число разнокачественных биологических категорий (и соответственно форм психического отражения), что эволюционный процесс развивался не линейно, а всегда несколькими параллельными путями. К тому же количество «экологических ниш» на земле необозримо, а ведь именно экологическими факторами, условиями жизни видов, занявших эти «ниши», определяются в первую очередь особенности их психической активности, а не их филогенетическим положением. Поэтому число «типичных представителей», которых необходимо изучать зоопсихологу, огромно (не говоря уже о том, что исследовать поведение всех животных вообще невозможно: число зоологических видов исчисляется миллионами, но даже, например, одних грызунов насчитывается свыше полутора тысяч видов!).
Отсюда следует, что первостепенная задача зоопсихологии состоит в установлении прежде всего самых общих закономерностей и основных этапов развития психики. Надежную основу для решения этой задачи представляет собой разрабатываемая советскими психологами теория деятельности. В этом плане большой вклад в разработку общих вопросов психического отражения у животных внес Алексей Николаевич Леонтьев. Мы имеем в виду прежде всего известные работы А.Н. Леонтьева «Проблема возникновения ощущения» и «Очерк развития психики», впервые опубликованные в 1940–1947 гг. и вошедшие затем в его труд «Проблемы развития психики», а также посмертно опубликованную работу «Психология образа». В последней он, в частности, указал, что многие проблемы зоопсихологии могут быть успешно разрешены, если рассматривать приспособление животных к жизни в окружающем их мире как приспособление к его дискретности, к связям наполняющих его вещей, их изменениям в пространстве. Напомним, что в «Очерке развития психики» А.Н. Леонтьев излагает свою концепцию стадиального развития психики в процессе эволюции животного мира.
Развивая свои идеи, А.Н. Леонтьев исходил из того, что не только у человека, но и у животных психика «включена» во внешнюю деятельность и зависит от нее, что «всякое отражение формируется в процессе деятельности животного».* Он подчеркивал, что психический образ является продуктом, практически связывающим субъект с предметной действительностью, что отражение животными среды находится в единстве с их деятельностью, что психическое отражение предметного мира порождается не непосредственными воздействиями, а теми процессами, с помощью которых животное вступает в практические контакты с предметным миром: первично деятельностью управляет сам предмет и лишь вторично его образ как субъективный продукт деятельности. По А.Н. Леонтьеву, возникновение и развитие психики обусловлено тем, «что выделяются процессы внешней деятельности, опосредующие отношения организмов к тем свойствам среды, от которых зависит сохранение и развитие их жизни».** На каждом новом этапе эволюции поведения и психики животных возникает все более полная подчиненность эффекторных процессов деятельности объективным связям и отношениям свойств предметов, во взаимодействие с которыми вступает животное. Предметный мир как бы все более «втягивается» в деятельность. Именно от характера этих связей, по мнению А.Н. Леонтьева, зависит, «будет ли отражаться и насколько точно будет отражаться в ощущениях животного воздействующее на него свойство предмета».*** При этом «материальную основу развития деятельности и чувствительности животных составляет развитие их анатомической организации»,**** т.е. морфологические структуры, в частности, нервная система, развивается вместе с деятельностью.
* Леонтьев А.Н. Проблемы развития психики. 3-е изд. М., 1972. С. 162.
** Там же. С. 159.
*** Там же. С.212.
**** Там же. С. 163.
Напрашивается вывод о том, что деятельность животного является источником познавательных способностей животных и что «познание мира» происходит у животных только в процессе и в итоге активного воздействия на окружающую среду, т.е. в ходе осуществления поведенческих актов. Чем более развиты, следовательно, двигательные возможности животного, тем выше и его познавательные способности. Можно поэтому сказать, что уровень психического отражения у тех или иных животных зависит от того, в какой мере они способны оказать воздействие на компоненты среды, насколько разнообразны и глубоки эти воздействия, а это в конечном итоге зависит от развития их двигательного аппарата. В этом мы видим воплощение теории деятельности в зоопсихологии и значение этой теории для успешного решения вопросов развития психики животных в процессе филогенеза (равно как и онтогенеза).
Как подчёркивал А.Н. Леонтьев, при изучении происхождения и развития психики необходимо учесть, что материальным субстратом психического отражения не обязательно является нервная система. Неудовлетворительность «нейропсихизма», согласно которому возникновение психики связано с появлением нервной системы, по А.Н. Леонтьеву, «заключается в произвольности допущения прямой связи между появлением психики и появлением нервной системы, в неучете того, что орган и функция, хотя и являются неразрывно взаимосвязанными, но вместе с тем связь их не является неподвижной, однозначной, раз и навсегда зафиксированной, так что аналогичные функции могут осуществляться различными органами».* Поэтому нельзя ставить возникновение психики «в прямую и однозначную связь с возникновением нервной системы, хотя на последующих этапах развития эта связь не вызывает, конечно, никакого сомнения».* Таким образом, Леонтьев подошел к проблеме происхождения и развития психики с глубоко осознанным пониманием диалектического характера этого процесса: от наиболее примитивных, зачаточных форм, субстратом которых является менее дифференцированная материя – протоплазма, к высшим формам психического отражения, возникающим на основе все более дифференцированной нервной ткани. Напрашивается вывод, что переход от, можно, очевидно, сказать, «плазматического» психического отражения к «нервному», связанный с появлением тканевой структуры у животных организмов, является подлинным диалектическим скачком, ароморфозом в эволюции животного мира. Нет нужды доказывать, что такой диалектико-материалистический подход к эволюции психики в корне отличается от плоско-эволюционистического, по существу, метафизического подхода, при котором усложнение поведения и психики животных однозначно связывается ТОЛЬКО (и притом часто произвольно) с изменениями в строении нервной системы.
* Там же. С. 10.
** Там же. С. 11.
Итак, намеченный А.Н. Леонтьевым подход к проблеме зарождения и развития психики приводит к выводу, что наличие нервной системы не является исходным условием развития психики, что, следовательно, психическая деятельность появилась раньше нервной деятельности. Очевидно, последняя возникла на таком уровне жизнедеятельности, когда осуществление жизненных функций стало уже настолько сложным, что возникла необходимость в таком специальном регуляторном аппарате, каковым и является нервная система. Затем же, уже в качестве органа психического отражения, нервная система (точнее, высшие отделы центральной нервной системы) стала необходимой основой и предпосылкой для дальнейшего развития психики. Этот сложный вопрос требует, конечно, специального рассмотрения, которое вышло за рамки этой статьи. Отметим лишь, что формирование нервной системы определялось решающими функциональными изменениями (коренными изменениями в жизнедеятельности организмов), т.е. имел место примат функции перед формой.
Важно отметить и указание А. Н. Леонтьева на то, что «если бы не существовало перехода животных к более сложным формам жизни, то не существовало бы и психики, ибо психика есть именно продукт усложнения жизни»,* что характер психического отражения «зависит от объективного строения деятельности животных, практически связывающей животное с окружающим его миром. Отвечая изменению условий существования, деятельность животных имеет свое строение, свою, так сказать, "анатомию". Это и создает необходимость такого изменения органов и их функций, которое приводит к возникновению более высокой формы психического отражения».** Как мы видим, A. Н. Леонтьев отчетливо отстаивал положение о примате функции в эволюции психики: изменившиеся условия адаптации к окружающей среде обусловливают изменение строения деятельности животных, вследствие чего меняется строение органов и их функционирование, что в свою очередь приводит к прогрессивному развитию психического отражения. Другими словами, А.Н.Леонтьев положил в основу своей концепции развития психики положение о том, что сущность этого процесса, его первопричина и движущая сила есть взаимодействие, представляющее собой материальный жизненный процесс, процесс установления связей между организмом и средой. Тем самым А. Н. Леонтьев распространил на сферу психического отражения известное положение марксистской философии о том, что всякое свойство раскрывает себя в определенной форме взаимодействия.
* Там же. С. 26.
** Там же.
В разработанной А. Н. Леонтьевым периодизации развития психики, охватывающей весь процесс эволюции животного мира, выделяются стадии элементарной сенсорной психики, перцептивной психики и интеллекта. Это было тем более знаменательно, что, за исключением В.А. Вагнера, усилия советских зоопсихологов (Н.Ю.Войтониса, Н. Н. Ладыгиной-Котс и др.) были направлены на изучение психики обезьян и не касались более низких уровней эволюции психики. Концепция стадиального развития психики, предложенная А. Н. Леонтьевым, являлась поэтому новым словом в науке, особенно на фоне традиционно упрощенного и сейчас уже неприемлемого деления на три якобы последовательные «ступени» – инстинкт, навык и интеллект. Концепция А.Н.Леонтьева принципиально отличается от такого понимания сущности эволюции поведения и психики прежде всего тем, что в ее основу положен не формальный признак деления всего поведения животных на врожденное (якобы «низшее») и благоприобретаемое («высшее»). Сейчас уже хорошо известно, что врожденное, инстинктивное поведение не является более примитивным по сравнению с индивидуально приобретаемым (научением), что, наоборот, оба этих компонента эволюционировали совместно, представляя единство целостного поведения, и что соответственно на высших этапах филогенеза мощное прогрессивное развитие получили как процессы научения, так и инстинктивные (генетически фиксированные) компоненты поведения животных.
К сожалению, однако, упомянутые упрощенческие, по существу метафизические, представления об этапах эволюции поведения животных, соответствующие уровню знаний 20-х годов, подчас преподносятся и поныне, особенно в работах некоторых физиологов. Так, например, В.Детьер и Э.Стеллар предлагают даже неизвестно по каким количественным данным вычерченное изображение кривых, из которых явствует, что простейшие сильны таксисами, у «простых» (?) многоклеточных же появляется, причем сразу «на полной мощности», рефлекс, насекомые являются всецело «инстинктивными» и «рефлекторными» животными, рыбы в основном также, но немного способны к обучению, птицы – наполовину «инстинктивными», наполовину «обучающимися», а млекопитающие – вполне обучающимися. При этом одна категория якобы сменяет другую: таксисы будто окончательно исчезают у птиц и низших млекопитающих, рефлексы и инстинкты теряют всякое приспособительное значение у низших приматов, которые якобы, как и низшие млекопитающие, адаптируются с помощью обучения и (приматы) отчасти – мышления. Все это всецело противоречит современным знаниям о поведении животных. Можно, например, напомнить, что такситы, эти ориентирующие компоненты любого поведенческого акта, непременно присущи поведению всех без исключения животных, а также человеку.
Проблемы психики животных не терпят упрощенчества, а тем более «глобальных» псевдорешений и дилетантского отношения, которые подчас встречаются в публикациях, авторы которых далеки от зоопсихологии, не считают нужным принять ее во внимание, но спешат выступить с поверхностными, нередко надуманными сенсационными «объяснениями». Только зоопсихологии, которая прежде всего является наукой об эволюции психики, по плечу задача научного познания закономерностей психики на всех уровнях филогенеза, вплоть до подступов зарождения человеческого сознания.
Иногда в качестве решающего критерия эволюции поведения и психики животных пытаются использовать степень сложности строения центральной нервной системы. В этих случаях за исходное принимается морфологическая структура, якобы определяющая филогенетический уровень и специфику поведения. Разумеется, при решении вопросов эволюции психики непременно надо учитывать строение и функции нервной системы изучаемых животных (наряду с другими их морфофункциональными признаками, например, двигательного аппарата). Но, как мы уже неоднократно указывали, особенности макростроения центральной нервной системы, особенно головного мозга, далеко не всегда отвечают особенностям поведения животных, уровню их психической деятельности. Достаточно указать, например, на птиц, психическая деятельность которых по уровню своего развития может быть приравнена к таковой у млекопитающих, в то время как головной мозг у птиц лишен вторичного мозгового свода – серой коры больших полушарий (неопаллидума), содержащей высшие ассоциативные центры. Аналогично поражает сложностью своего поведения, граничащего подчас с интеллектуальным, крыса, мозг которой является весьма примитивным по своему строению: с гладкой поверхностью больших полушарий, лишенной борозд (последние появляются среди грызунов и зайцеобразных только у бобра, сурка и зайцев).
Тем не менее, нередко высказывается мнение, будто прогресс в эволюции поведения животных первично определяется морфологическими изменениями в центральной нервной системе (например, увеличением числа нейронов в результате мутации). Совершенно ясно, что единственно верным является противоположный подход, при котором признается примат функции перед формой (строением), подход, успешно осуществляемый и в советской эволюционной морфологии. При решении вопросов развития психики (как в фило-, так и в онтогенезе) и вообще в любых зоопсихологических оценках это означает, что появляются вторично в результате изменения поведения морфологические изменения, которые в свою очередь порождаются изменениями в образе жизни, когда возникает необходимость в установлении новых связей с компонентами окружающей среды. А поскольку поведение представляет собой совокупность всех двигательных актов животного, то можно сказать, что связанные с эволюцией психики морфологические изменения были первично обусловлены изменениями в двигательной активности животного, биологическое значение которой и состоит в установлении жизненно необходимых для организма связей со средой. Конечно, надо при этом помнить, что мы имеем здесь дело с диалектическим процессом, ибо затем возникшие новые формы (морфологические образования или количественные изменения) уже в свою очередь оказывают направляющее, развивающее влияние на функцию (поведение). Но во всяком случае прогрессивные изменения в нервной (и рецепторной) системе возникают как следствие адаптивных изменений двигательной активности животного и сопряженных с ними адекватных изменений в строении двигательного аппарата. Вот где следует искать первоисточник эволюционных преобразований в психической сфере животных.
* * *
Со времени опубликования А.Н. Леонтьевым концепции стадиального развития психики в процессе эволюции прошло более сорока лет, и можно с уверенностью сказать, что в целом эта концепция выдержала «проверку временем» и не утратила своего научного значения как общая теория эволюции психики в мире животных. Но это сорокалетие было периодом поистине бурного развития науки о поведении животных, и за эти годы был накоплен огромный материал, заставляющий нас во многом поновому посмотреть на психическую деятельность животных и внести соответствующие поправки в прежние представления, в частности касающиеся оценки уровня психического развития того или иного систематического таксона животных. Пришлось нам внести существенные уточнения в предложенную А.Н. Леонтьевым периодизацию эволюции психики.
Прежде всего оказалось необходимым ввести в эту периодизацию новые подразделения – уровни психического развития. В пределах как элементарной сенсорной, так и перцептивной психики мы выделили низший и высший уровни развития, имея при этом в виду, что в дальнейшем следует ввести еще дополнительно промежуточные уровни. Эти уровни обозначают, по существу, процесс развития самой элементарной или соответственно перцептивной психики.
Исходя из общего положения, что «каждая новая ступень психического развития имеет в своей основе переход к новым внешним условиям существования животных и новый шаг в усложнении их физической организации»,* А.Н. Леонтьев называет в качестве общего критерия СТАДИЮ ЭЛЕМЕНТАРНОЙ СЕНСОРНОЙ ПСИХИКИ, обусловленную приспособлением к вещно оформленной среде, способность к отражению отдельных свойств среды. При этом двигательная активность, деятельность, по Леонтьеву, «пробуждается тем или иным воздействующим на животное свойством, на которое она вместе с тем направлена, но которое не совпадает с теми свойствами, от которых непосредственно зависит жизнь данного животного».**
* Там же. С.193.
** Там же. С. 159.
Выделенный нами низший уровень стадии элементарной сенсорной психики характеризуется наиболее примитивными, точнее сказать, зачаточными проявлениями психического отражения, когда животные (простейшие, многие низшие многоклеточные) реагируют на биологически значимые свойства компонентов среды на сигналы о присутствии жизненно важных условий среды, как, хотя и четко, но еще очень несовершенным образом. Эти реакции еще не носят характера подлинного активного ориентировочного поведения. В основном это – изменение скорости и направления движения (только в водной среде) в градиенте раздражителя особой модальности (химического, температурного и т.п.), но не активный поиск благоприятных условий среды и жизненно необходимых ее компонентов. К тому же сигнальное значение свойств последних в большей степени еще слито с непосредственно жизненно необходимыми свойствами. По этой причине и отсутствует подлинная ориентация: нет еще топотаксисов, а представлены лишь первичные элементы таксисного (ориентировочного) поведения – ортотакситы и клинотаксисы, а также фобические реакции, позволяющие животному избегать неблагоприятных внешних условий. Двигательная активность представлена на этом низшем уровне развития психики самыми элементарными формами локомоции – кинезами, осуществляемыми в основном (за исключением амеб и споровиков) с помощью наиболее примитивного, первичного в животном мире двигательного аппарата – жгутиков, у более высокоорганизованных – при помощи ресничек. Вместе с тем, особенно у многоклеточных животных, уже появляются сократительные движения, лежащие в основе поведения всех высших животных. Манипуляционные движения (активное перемещение предметов, чаще всего в сочетании с физическим воздействием на них) ограничиваются у одноклеточных созданием потоков воды в окружающем их пространстве, у примитивных многоклеточных же они уже представлены простыми формами перемещения объектов с помощью щупалец, этих первых специальных эффекторов, т.е. органов манипулирования.
Пластичность поведения на низшем уровне элементарной сенсорной психики еще очень невелика. Выражается это, в частности, в том, что индивидуальная изменчивость поведения ограничена элементарной формой научения – привыканием, и лишь в отдельных случаях, возможно, встречаются зачатки ассоциативного научения. Особенно это относится к одноклеточным, которым подчас вовсе отказывают в способности к (ассоциативному) научению. Во всяком случае можно с уверенностью сказать, что на этом, начальном, уровне развития психики не может быть сколько-нибудь выраженного ассоциативного научения ввиду того, что еще не совсем сформировалась способность животных к реагированию на «биологически нейтральные», т.е. лишенные жизненного значения, свойства компонентов среды как на сигналы. А без этого, понятно, невозможно образование временных связей. В целом напрашивается вывод, что на низшем уровне элементарной сенсорной психики психическое отражение еще не приобрело самостоятельного значения, что наряду с психическим отражением внешняя активность животных в большей мере еще детерминируется также элементами допсихического отражения, которые составляют своеобразное единство с элементами зарождающегося психического отражения. Конкретно это означает, что на этом уровне представлены только элементы чувствительности, ее зачатки в самом примитивном виде, сосуществующие в поведении этих животных с ярко выраженной раздражимостью. Очевидно, только на более высоких уровнях эволюции значение раздражимости как детерминанты поведения сводится на нет.
Этот вывод относится прежде всего к простейшим. Но при этом следует учесть, что простейшие – особая, рано отклонившаяся филогенетическая ветвь, и что главное, современные простейшие являются продуктом эволюционного процесса, длившегося 1–1,5 миллиарда лет! Поэтому и строение и поведение современных «простейших» являются уже намного более сложными, чем у их ископаемых предков, и к тому же четко специализированными. Есть поэтому все основания полагать, что на заре зарождения животной жизни должен был существовать еще более низкий, чем у современных одноклеточных, действительно первоначальный уровень психического отражения, который давно уже исчез вместе с ископаемыми животными и о характере которого мы можем строить лишь гипотетические предположения. Но правомерно допустить, что на том исходном уровне развития психического отражения первичные формы внешней двигательной активности, поведения детерминировались только раздражимостью.
Высший уровень стадии элементарной сенсорной психики мы определяем как такой этап, на котором психическое отражение хотя также еще полностью и отвечает сформулированным А.Н. Леонтьевым общим признакам данной стадии, т.е. ограничено рамками ощущений, но сочетается с достаточно сложным поведением. Находящиеся на этом уровне животные – высшие (кольчатые) черви, брюхоногие моллюски (улитки) и некоторые другие беспозвоночные – не только уже четко отличают сигнальное значение «биологически нейтральных» раздражителей, но и обладают уже развитым таксисным поведением: на этом уровне впервые появляются высшие таксисы (топотаксисы), с помощью которых животные способны не только избегать неблагоприятных условий среды и уходить от них, но и вести активный поиск положительных раздражителей, приводящий их к жизненно необходимым компонентам среды не случайно, а «целенаправленно». Возможность точно локализировать биологически значимые объекты, активно взаимодействовать с ними экономит много энергии и времени при осуществлении поведенческих актов. Все это подняло жизнедеятельность на качественно иной уровень и существенно обогатило психическую сферу этих животных.
Основу такого эволюционного новоприобретения составляет развитая двигательная активность, которая проявляется в весьма разнообразных формах. Особенно это относится к локомоторным движениям, которые представлены ползанием, рытьем в грунте, плаванием, причем впервые осуществлен выход из воды на сушу (дождевые черви, наземные пиявки и улитки), т.е. в совершенно иную среду обитания с в корне отличными условиями существования. Впервые появился важнейший для дальнейшей эволюции психики орган манипулирования – челюсти, с помощью которых выполняются довольно разнообразные манипуляционные движения, но конечности представлены лишь в примитивнейшем, зачаточном виде, и то лишь у некоторых червей (параподии полихет). Соответственно и локомоторные движения выполняются медленно: за отдельными исключениями животные неспособны на этом уровне эволюции психики развить большую скорость. Поведение характеризуется еще низкой пластичностью, индивидуальный опыт, процессы изучения играют в жизни этих животных еще небольшую роль, решающее значение имеют жесткие врожденные программы поведения. Но все же на этом уровне уже вполне проявляется способность к образованию подлинных ассоциативных связей, хотя формируются они, как правило, в ограниченных пределах, медленно и на основе лишь большого числа сочетаний, а сохраняются недолго. Обнаруживаются и некоторые зачатки сложных форм поведения, характерные для представителей стадии перцептивной психики (конструктивной деятельности, территориальности, антагонистического поведения, общения).
В целом психика животных, представляющих высший уровень элементарной сенсорной психики, предстает перед нами в достаточно пестром виде: с одной стороны, эти животные, так же как наиболее низкоорганизованные представители животного мира, ориентируют свое поведение только по отдельным свойствам предметов (или их сочетаниям), и еще отсутствует способность к восприятию предметов как таковых (лишь у некоторых хорошо и быстро плавающих хищных многощетинковых червей да у некоторых улиток, может быть, существует какое-то элементарное предметное восприятие); с другой стороны, наличие более совершенных двигательных и сенсорных систем, особенно же нервной системы типа ганглиарной лестницы и головы с мозговым ганглием, позволяет животным на этом уровне психического развития устанавливать весьма разнообразные и подчас довольно сложные связи с компонентами окружающей среды, в результате чего достигается более многостороннее и более содержательное психическое отражение этих компонентов.
* * *
Стадия перцептивной психики характеризуется А. Н. Леонтьевым прежде всего «способностью отражения внешней объективной действительности уже не в форме объективных элементарных ощущений, вызываемых отдельными свойствами или их совокупностью, но в форме отражения ВЕЩЕЙ»*, причем переход к этой стадии связан с изменением строения деятельности животных, которое «заключается в том, что уже наметившееся ранее содержание ее, объективно относящееся не к самому предмету, на который направлена деятельность животного, но к тем условиям, в которых этот предмет объективно дан в среде, теперь выделяется».** И далее: «Если на стадии элементарной сенсорной психики дифференциация воздействующих свойств была связана с простым их объединением вокруг доминирующего раздражителя, то теперь впервые возникают процессы интеграции воздействующих свойств в единый целый образ, их объединение как свойств одной и той же вещи. Окружающая действительность отражается теперь животным в форме более или менее расчлененных образов отдельных вещей».*** В результате возникает и новая форма закрепления опыта животных – двигательные навыки, определяемая А. Н. Леонтьевым как закрепленные операции, которыми он обозначает «особый состав или сторону деятельности, отвечающую условиям, в которых дан побуждающий ее предмет».****
* Там же. С. 176.
** Там же.
*** Там же. С.177.
**** Там же.
Применяя эти критерии с учетом современных знаний о поведении животных, нам пришлось значительно понизить филогенетическую грань между элементарной сенсорной и перцептивной психикой по сравнению с представлениями А. Н. Леонтьева, который провел эту грань «выше» рыб. На самом деле, как сегодня хорошо известно, указанные А. Н. Леонтьевым критерии перцептивной психики вполне соответствуют психической деятельности рыб и других низших позвоночных и даже, правда с определенными ограничениями, высших беспозвоночных. Уже по этой причине нельзя согласиться с высказанным А. Н. Леонтьевым мнением, будто появление перцептивной психики обусловлено переходом животных к наземному образу жизни.
Высшие беспозвоночные (членистоногие и головоногие моллюски) достигли НИЗШЕГО УРОВНЯ перцептивной психики и соответственно проявляют в своем поведении еще немало примитивных черт. Поведение этих животных, составляющих подавляющее большинство всех существующих на земле видов, еще очень мало изучено, и не исключено, что среди низших членистоногих многие виды остались за пределами перцептивной психики. Но и у высших членистоногих, в том числе и у наиболее высокоорганизованных насекомых, в поведении преобладают ригидные, «жестко запрограммированные» компоненты, а пространственная ориентация осуществляется по-прежнему преимущественно по отдельным свойствам предметов. Предметное же восприятие, хотя уже представлено в развитой форме, играет в поведении членистоногих лишь подчиненную роль (значительно большее значение оно имеет, очевидно, для головоногих моллюсков). Но, с другой стороны, ведь и у позвоночных (включая высших) пространственно-временная ориентация осуществляется преимущественно на уровне элементарных ощущений (ольфакторных, акустических, оптических и др.). Достаточно вспомнить, что именно на этом механизме построено реагирование на ключевые раздражители, детерминирующие жизненно необходимые действия как для низших, так и для высокоорганизованных животных.
Представители низшего уровня перцептивной психики обитают повсеместно, во всех климатических зонах, во всех средах, во всех «экологических нишах». Локомоторные способности этих животных проявляются в очень разнообразных и весьма сложных формах. В сущности, здесь представлены все виды локомоции, которые вообще существуют в мире животных: разнообразные формы плавания, в том числе реактивное (у головоногих моллюсков), ныряние, ползание (в воде и на суше), ходьба, бег, прыгание, лазанье, передвижение с помощью цепляния и подтягивания тела (например, у осьминогов), роющее или грызущее передвижение в субстрате (в грунте, в древесине и т.п.) и др. Здесь же мы встречаем тех животных, которые впервые освоили воздушное пространство, что первые летающие животные на земле (насекомые), причем полет представлен у них в совершенстве. Помимо этой активной формы (с помощью крыльев) существует и пассивная (с помощью паутинок). Возникновение такой качественно новой формы локомоции, как полет, было, конечно, важным событием в эволюции животного мира. Значительно большее, однако, значение имело для эволюции психики появление на этом уровне подлинных, сложно устроенных конечностей – ног, в результате чего не только существенно расширилась сфера активности животных, но и возникли совершенно новые, принципиально иные условия для активного воздействия на компоненты среды, возникли предпосылки полноценного манипулирования предметами. Конечно, на данном эволюционном уровне манипулирование еще слабо развито, хотя на этом уровне впервые появляются специальные хватательные конечности (клешневидные педипальцы скорпионов и лжескорпионов, клешни раков и крабов, хватательные конечности богомолов и т.п.). Однако главным органом манипулирования остается по-прежнему челюстной аппарат, который у членистоногих весьма сложно устроен. Тем не менее челюсти, как и хватательные конечности членистоногих, производят только весьма однообразные (именно «клешневидные») движения. Иногда в воздействиях на предметы участвуют и ходильные конечности (креветки, пауки, жук-скарабей и др.). Дифференцированные и разнообразные манипуляционные движения же у членистоногих отсутствуют. У головоногих, правда, манипуляционная активность играет значительно большую роль, и они снабжены превосходными хватательными органами – щупальцами, которые нередко справедливо называют руками, функциональными аналогами которых они действительно являются. Поэтому головоногие (особенно осьминоги) способны, вероятно, к значительно более разностороннему и полноценному двигательному обследованию объектов манипулирования, чем членистоногие, тем более что головоногие обладают превосходным зрением, аналогичным таковому позвоночных, и движения щупалец происходят в поле их зрения. Однако поведение головоногих моллюсков еще крайне слабо изучено.
Изучение форм и уровня развития локомоции и манипулирования имеет, несомненно, исключительное значение для познания психических способностей животных. Мы исходим при этом из того, что психические процессы всегда воплощаются или в локомоторной, или в манипуляционной внешней активности (а также в некоторых особых формах демонстрационного поведения), а первично зависят от уровня развития и степени дифференцированности этих движений. Это относится ко всем формам и возможностям познавательной деятельности животных, вплоть до высших психических способностей; всегда любая встающая перед животным задача может быть решена только или локомоторным, или манипуляционным путем. У членистоногих абсолютно доминирует локомоторное решение задач. Соответственно преобладает и пространственная ориентация над манипуляционным обследованием предметов. Последнее, вероятно, вообще встречается, причем лишь в самых примитивных формах, среди членистоногих только у некоторых высших насекомых, пауков и некоторых высших ракообразных.
Пространственная ориентация характеризуется на низшем уровне перцептивной психики четко выраженным активным поиском положительных раздражителей (наряду с мощно и многообразно развитым защитным поведением, т.е. избеганием отрицательных раздражителей). Большую роль играют в жизни этих животных и впервые появившиеся в филогенезе мнемотаксисы (ориентация по индивидуально выученным ориентирам). Процессы научения занимают в поведении животных на этом уровне вообще заметное место, но примитивной чертой является их неравномерное распределение по разным сферам жизнедеятельности (преимущественно способность к научению проявляется в пространственной ориентации и пищедобывательной деятельности), а также подчиненное положение научения по отношению к инстинктивному поведению: научение служит здесь прежде всего для совершенствования врожденных компонентов поведения, для придания им должной пластичности, но оно еще не приобрело самостоятельного значения, как это имеет место у представителей высшего уровня перцептивной психики.
Здесь необходимо, однако, вновь сделать оговорку относительно головоногих моллюсков, которых, вероятно, вообще следует поместить на более высокий уровень, чем членистоногих, тем более что они по многим признакам строения и поведения проявляют черты аналогии с позвоночными, а также сопоставимы с последними по размерам. Наряду с высокоразвитыми формами инстинктивного поведения (территориальное и групповое поведение, ритуализация, сложные формы ухода за потомством – икрой), которые, правда, встречаются и у членистоногих, у головоногих описаны проявления «любопытства» по отношению к «биологически нейтральным» объектам и высокоразвитые конструктивные способности (сооружение с помощью «рук» валов и построек-убежищ у осьминога). Головоногие (осьминоги) в отличие от членистоногих (включая «одомашненных» пчел) способны общаться с человеком (это первый случай на филогенетической лестнице) и поэтому могут даже приручаться!
* * *
Обозначенные выше характеристики дают достаточное представление о том, что низший уровень, точнее, низшие уровни стадий перцептивный психики обнаруживают еще ряд примитивных признаков, унаследованных от элементарной сенсорной психики. Безусловно, существуют и промежуточные уровни перцептивной психики, которые еще предстоит выделить в ряду позвоночных. Здесь мы вкратце коснемся только высшего уровня перцептивной психики. На этом уровне находятся высшие позвоночные (птицы и млекопитающие), к которым А.Н. Леонтьев и относил всю стадию перцептивной психики. Поведение этих животных хорошо изучено. Мы имеем здесь дело с вершиной эволюции психики, с высшими проявлениями психической деятельности животных. Это относится как к двигательной, так и к сенсорной сферам как к компонентам как врожденного, так и приобретенного поведения. Иными словами, здесь достигают наивысшего развития как инстинктивное поведение, так и способность к его индивидуальной модификации, т.е. способность к научению. О наивысших проявлениях этой способности мы говорим как об интеллектуальном поведении, основанном на процессах элементарного мышления. По меньшей мере на высшем уровне перцептивной психики у животных уже складываются определенные «образы мира». У животных следует, очевидно, понимать под психическим образом практический опыт их взаимодействия с окружающим миром, который актуализируется в результате повторного восприятия его конкретных предметных ситуаций.
О характере этих образов можно судить по результатам изучения ориентировочно-исследовательской деятельности млекопитающих, осуществленного на серых крысах в нашей лаборатории. Так, например, было установлено, что в ходе активного ознакомления с особенностями нового пространства или нового предмета у животных наблюдается своеобразный процесс уподобления внешний активности, поведения особенностям обследуемого пространства или предмета. При этом происходит постепенное увеличение степени адекватности поведения животных условиям нового пространственного окружения или свойствам предмета. Вместе с тем ориентировочно-исследовательская деятельность всегда разворачивается под определяющим влиянием формирующегося образа, который обусловливает возможность дальнейшего обследования пространства или предмета. По мере того как возрастает адекватность поведения в новой ситуации, его соответствие объективным условиям этой ситуации, ориентировочно-исследовательская деятельность угасает и животное возвращается к повседневной жизнедеятельности. Это позволяет говорить о том, что к этому времени образ данной ситуации и действий животного в ней уже сформирован. Эта приспособленность поведения к новым условиям окружающей среды и является биологически адекватным, необходимым для выживания результатом формирования «образов мира» у животных. Проведенное исследование является первой попыткой подойти со стороны зоопсихологии к конкретизации и анализу процесса формирования образа.
Надо думать, что образы будут существенно отличаться друг от друга в зависимости от того, на какой основе они формировались. Так, образы, возникшие на основе лишь локомоторной активности (при ознакомлении с новым пространством), будут иными, чем те, которые формировались на основе манипуляционных действий (при манипуляционном обследовании новых предметов). Локомоция дает животному обширные пространственные представления, манипулирование же – углубленные сведения о физических качествах и структуре предметов. Оно позволяет полноценно выделять предметные компоненты среды как качественно обособленные самостоятельные единицы и подвергнуть их такому обследованию, которое у высших представителей животного мира служит основой интеллекта. Локомоторные формы ориентации и реагирования на ситуации новизны для этого недостаточны.
* * *
А.Н. Леонтьев выделил особую (третью) «стадию интеллекта», причем специально для человекообразных обезьян. Главный критерий интеллектуального поведения, по Леонтьеву, – перенос решения задачи в другие условия, лишь сходные с теми, в которых оно впервые возникло, и объединение в единую деятельность двух отдельных операций (решение «двухфазных» задач). При этом он указывал на то, что сами по себе формирование операции и ее перенос в новые условия деятельности «не могут служить отличительными признаками поведения высших обезьян, так как оба эти момента свойственны также животным, стоящим на более низкой стадии развития. Оба эти момента мы наблюдаем, хотя в менее яркой форме, также и у многих других животных – у млекопитающих, у птиц».* Но от последних операции человекообразных обезьян отличаются особым качеством – двухфазностью, причем первая, подготовительная фаза вне связи со следующей фазой (фазой осуществления) лишена какого бы то ни было биологического смысла. «Наличие фазы подготовления и составляет характерную черту интеллектуального поведения. Интеллект возникает, следовательно, впервые там, где возникает процесс подготовления возможности осуществить ту или иную операцию или навык».**
* Там же С. 189.
** Там же. С. 191.
Следует отметить, что сама по себе двухфазность, наличие подготовительной и завершающей фаз, как мы сегодня знаем, присуща любому поведенческому акту, и, следовательно, в такой общей формулировке этот признак был бы недостаточен как критерий интеллектуального поведения животных. Однако при решении задач на уровне интеллектуального поведения, как подчеркивает А.Н. Леонтьев, «существенным признаком двухфазной деятельности является то, что новые условия вызывают у животного уже не просто пробующие движения, но пробы различных прежде выработавшихся способов, операций».* Отсюда вытекает и важнейшая для интеллектуального поведения способность «решать одну и ту же задачу многими способами»,** что в свою очередь доказывает, что здесь «операция перестает быть неподвижно связанной с деятельностью, отвечающей определенной задаче, и для своего переноса не требует, чтобы новая задача была непосредственно сходной с прежней».** В итоге при интеллектуальной деятельности «возникает отражение не только отдельных вещей, но и их отношений (ситуации) ...
Эти обобщения животного, конечно, формируются так же, как и обобщенное отражение им вещей, т.е. в самом процессе деятельности».****
* Там же.
** Там же.
*** Там же.
**** Там же. С.192.
Общая характеристика и критерий интеллекта животных, предложенные А.Н. Леонтьевым, сохраняют свое значение и на сегодняшний день. Однако следует иметь в виду, что Леонтьев проанализировал эту сложнейшую проблему традиционно только на примере лабораторного изучения способности шимпанзе к решению искусно придуманных экспериментатором задач с помощью орудий. Вместе с тем, как мы сейчас знаем, двухфазная орудийная деятельность в весьма разнообразных формах распространена и среди других животных, больше всего даже среди птиц, и, как показывают данные ряда современных авторов, может быть с не меньшим успехом воспроизведена у них в эксперименте. Каждый год приносит новые неожиданные данные, свидетельствующие о том, что антропоиды не обладают монополией на решение таких задач. К тому же орудийная деятельность – не обязательный компонент интеллектуального поведения, которое, как также свидетельствуют современные данные, может проявляться и в других формах, причем опять же не только у антропоидов, но и у разных других животных (вероятно, даже у голубей). А с другой стороны, орудийные действия встречаются и у беспозвоночных, у которых явно не может быть речи об интеллектуальных формах поведения. Наконец, практически невозможно провести четкую грань между разнообразнейшими сложными навыками и интеллектуальными действиями высших позвоночных (например, у крыс), поскольку в ряду позвоночных навыки, постепенно усложняясь, плавно переходят в интеллектуальные действия.
Таким образом, в том или ином виде, часто в более элементарных формах, интеллектуальное поведение (или трудноотделимые от него сложные навыки) довольно широко распространено среди высших позвоночных. Поэтому если исходить из критериев А. Н. Леонтьева, то в соответствии с современными знаниями невозможно провести грань между некоей особой стадией интеллекта и якобы ниже расположенной стадией перцептивной психики. Все говорит за то, что способность к выполнению действий интеллектуального типа является одним из критериев высшего уровня перцептивной психики, но встречается эта способность не у всех представителей этого уровня, не в одинаковой степени и не в одинаковых формах.
Итак, приведенные А. Н. Леонтьевым критерии интеллекта уже не применимы к одним лишь антропоидам, а «расплываются» в поведении и других высших позвоночных. Но означает ли это, что шимпанзе и другие человекообразные обезьяны утратили свое «акме», свою исключительность? Отнюдь нет. Это означает лишь, что не в этих общих признаках интеллектуального поведения отражаются качественные особенности поведения антропоидов. Решающее значение имеют в этом отношении качественные отличия манипуляционной активности обезьян, позволяющие им не только улавливать наглядно воспринимаемые связи между предметами, но и знакомиться со строением (в том числе и внутренним) объектов манипулирования. При этом движения рук и соответственно тактильно-кинестетические ощущения вполне сочетаются со зрением, из чего следует, что зрение у обезьян также «воспитано» мышечным чувством, как это показал для человека еще И.М.Сеченов. Вот почему обезьяны способны к значительно более глубокому и полноценному познанию, чем все другие животные. Эти особенности манипуляционно-исследовательской деятельности дают основание полагать, что, вопреки прежним представлениям, обезьяны (во всяком случае человекообразные) способны усмотреть и учесть причинно-следственные связи и отношения, но только наглядно воспринимаемые, «прощупываемые» физические (механические) связи. На этой основе и орудийная деятельность обезьян поднялась на качественно иной, высший уровень, как и вообще отмеченные отличительные особенности их манипуляционной активности привели к недосягаемому для других животных развитию психики, к вершине интеллекта животных. Главное при всем этом заключается в том, что развитие интеллекта (как и вообще психики) шло у обезьян в принципиально ином, чем у других животных, направлении – в том единственном направлении, которое только и могло привести к возникновению человека и человеческого сознания: прогрессивное развитие способности к решению манипуляционных задач на основе сложных форм предметной деятельности легло в основу того присущего только обезьянам «ручного мышления» (термин И. П. Павлова), которое в сочетании с высокоразвитыми формами компенсаторного манипулирования являлось важнейшим условием зарождения трудовой деятельности и специфически человеческого мышления. Сказанное дает, очевидно, основания для выделения в пределах перцептивной психики специального «наивысшего уровня», который хотя и будет представлен обезьянами, однако по указанным выше причинам не будет соответствовать «стадии интеллекта», как она была сформулирована А. Н. Леонтьевым.
Проблема эволюции психики – чрезвычайно сложная и требует еще всестороннего обстоятельного изучения. Советские зоопсихологи с большой благодарностью вспоминают Алексея Николаевича Леонтьева, который не только глубоко понимал значение зоопсихологических исследований, но и сделал очень много для утверждения и прогресса нашей науки.
Зоопсихология*
* Тенденции развития психологической науки / Под ред. Л. И. Анцыферовой, Б. Ф. Ломова. М., 1989. С. 208-221.
Зоопсихология – одна из основных базовых отраслей общей психологии. Ее предмет охватывает всю проблематику психической деятельности современных животных, эволюцию психики, т.е. весь фило- и онтогенез психики животных, биопсихологические предпосылки антропогенеза, предысторию и зарождение сознания человека, все биопсихологические аспекты развития психики у современного человека, а следовательно и существенную часть проблемы социального и биологического в психике человека. Ясно, что без учета развития психики, особенно в эволюционном плане, не может быть диалектико-материалистической психологии.
Объект зоопсихологии – поведение животных, которое со своих специфических позиций изучают также другие науки (этология, нейрофизиология, физиология ВНД, бионика и др.). Под поведением мы понимаем всю совокупность проявлений внешней, преимущественно двигательной активности животного, направленной на установление жизненно необходимых связей организма со средой. Зоопсихолог изучает поведение животных как первопричину психического отражения, точнее, весь комплекс проявлений поведения и психики, единый процесс психического отражения как продукт внешней активности животного. Иными словами, зоопсихолог никогда не ограничивается изучением лишь одного поведения без анализа его психического аспекта.
В этом заключается специфика зоопсихологического исследования в отличие, например, от этологического. Этология – раздел биологии, точнее, зоологии. Ее задача – изучение общебиологических основ и закономерностей поведения животных, поведения как экологического фактора адаптации в онто- и филогенезе. В созданных этологами учениях о «биологии поведения», в частности современной концепции инстинктивного поведения животных, центральное место занимают видотипичные, генетически фиксированные компоненты поведения. Чрезвычайно важным разделом зоопсихологии является сравнительная психология, призванная изучить общие закономерности и предпосылки возникновения человеческого сознания, выявить в онто- и филогенезе животных психические компоненты, свидетельствующие об общности происхождения психических процессов у животных и человека, равно как и качественные отличия человеческой психики.
Советская зоопсихология воплощает диалектико-материалистическую тенденцию в развитии зоопсихологии. Ее конкретно-научная методология – деятельностный подход, который открывает строго научный путь к познанию закономерностей психической деятельности животных.
Для более детального ознакомления с предметом зоопсихологии можно рекомендовать учебник зоопсихологии, допущенный Минвузом СССР в этом качестве для студентов университетов. О специфических методах исследования, истории, о положении зоопсихологии в системе наук, о ее новом концептуальном аппарате можно получить представление в названном источнике и ряде недавно опубликованных трудов автора.
Говоря об объекте зоопсихологических исследований – поведении животных, следует указать на то, что если «обычная» психология имеет дело лишь с одним, хотя и весьма непростым, объектом исследования, с одним лишь видом – человеком, то никто не может сказать, сколько их у зоопсихологии, ибо никто не знает, сколько существует видов животных на земле. Одних насекомых, по весьма осторожной оценке советского энтомолога Г.Я Бей-Биенко, один-полтора миллиона видов, тем более что ежегодно открывают тысячи (!) новых видов. Позвоночных, правда, значительно меньше – «всего» 40–45 тыс. видов. А ведь каждый вид имеет свои биологические, а следовательно и психические, особенности, и чтобы создать полноценную зоопсихологию, надо было бы как следует изучить хотя бы по одному представителю каждого рода или на худой конец семейства. Едва ли эта цель достижима, во всяком случае мы от нее бесконечно далеки: более или менее обстоятельно и достоверно изучено на сегодняшний день всего несколько видов млекопитающих, птиц, рыб и насекомых. Об остальных (включая, например, гигантский тип моллюсков – около 130 тыс. видов!) мы все еще почти ничего не знаем: имеются лишь кое-какие отрывочные сведения. Отсюда следует, что в психологии термин «животное» (или «животные») можно употребить лишь в сравнительно-психологическом понимании как синоним «животного мира» (обычно имея в виду высших его представителей), точнее, когда речь идет о дочеловеческом уровне развития психики в целом.
Одна из тенденций современной зоопсихологии состоит в специализации, выделении отдельных отраслей, задача которых состоит в планомерном накоплении конкретного материала, на основе которого можно было бы индуктивно строить выводы, общезначимые для крупных зоологических таксономических единиц. Необходимость создания таких новых подразделений диктуется также запросами практики. Примером могут служить первые шаги новой отрасли общей зоопсихологии, названной нами «ихтиоп-сихологией», т.е. психологией рыб (см. ниже).
Советские зоопсихологи ищут пути к познанию психики животных, ее эволюции и ее онтогенетического развития, прежде всего опираясь на детальный анализ двигательной активности как источника психического отражения у животных, исходя из того, что «познание мира» происходит у них только в процессе и в итоге воздействия на окружающую среду. Чем более развиты двигательные возможности животного, тем выше и его познавательные способности. Можно поэтому сказать, что уровень психического отражения у того или иного вида животных зависит от того, в какой мере он способен оказать воздействия на компонент среды, насколько разнообразны и глубоки эти воздействия, а это в конечном итоге зависит от развития его двигательного аппарата.
Эта позиция – творческая, не оставляющая места спекулятивным догадкам и «гипотезам» о сущности и эволюции психики. Это путь психологического анализа внешней активности, поведения животного в ходе решения задач, и об этом уже много писалось. Это путь изучения психики животных, базирующийся на диалектическом единстве поведения и психики; это научный поиск, основанный на объективном психологическом анализе структуры поведения животных, на учете экологических особенностей изучаемого вида и специфики психических функций на разных филогенетических уровнях, исходя из примата локомоторно-манипуляционной активности в процессе развития психики. Таковы исходные методологические основы концептуального аппарата новой советской зоопсихологии. Они же определяют тенденции ее дальнейшего развития.
* * *
Наиболее пагубное влияние всегда оказывало и, к сожалению, продолжает оказывать антропоморфическое толкование поведения животных, основанное на стирании качественных различий между поведением и психикой человека и животных. Антропоморфизм в зоопсихологии проявляется в субъективном толковании поведения животных по аналогии с психическими функциями и состояниями человека. «Отцом» антропоморфической зоопсихологии следует, очевидно, считать известного английского биолога Дж. Дж. Роменса, сподвижника Ч. Дарвина, внесшего немалый вклад в эволюционную теорию, но приписывавшего даже членистоногим и червям человеческие способности вплоть до разума.
Необходимо, однако, иметь в виду, что сто лет тому назад еще не была разработана зоопсихологическая терминология и употребляемым понятиям придавалось весьма нечеткое значение. Термин «разум» употребляется скорее как антоним понятия «инстинкт» для обозначения изменчивого поведения. Дарвин, которого очень интересовала проблема биологической изменчивости, в письме от 16 апреля 1881 г. писал Роменсу: «Я не пытался определить разум, но привел Ваши слова о значении опыта и показал, насколько они применимы к червям. По-моему, о них следует сказать, что они до некоторой степени руководствуются разумом, во всяком случае они не подчиняются слепому инстинкту... Хотя Фабр упорно настаивает на неизменяемости инстинктов, однако ясно, что некоторая изменяемость существует...». Эта цитата характеризует и позицию самого Дарвина, который был убежден в том, что человеческий разум «развился из такого же разума, каким обладают животные» (из письма Т. Г. Фарреру от 28 августа 1881 г.).
В наше время биологизация поведения человека проявляется, например, в том, что ему приписывается некая роковая «исконная биологическая агрессивность», якобы повинная во всех бедах человечества. Нам не раз приходилось реагировать на эту ложную, крайне реакционную версию. Напомним лишь, что ее нельзя признать научной хотя бы уже потому, что она выведена путем умозрительного сопоставления разнокачественных явлений, т.е. на основе аналогий (к тому же мнимых), а не гомологии.
Термин «агрессивность» в применении к животным, безусловно, неудачен, ибо сам по себе означает специфические человеческие действия и тем самым чреват социологизаторским пониманием обозначаемых этим термином форм поведения животных, особенно в сравнительно-психологическом плане (например, при сравнении конфронтации в мире животных с войнами), и выхолащиванием его общественно-исторически обусловленного содержания. Однако он стал уже неотъемлемой частью этологической и зоопсихологической терминологии, а к тому же он отчасти употребляется и для обозначения некоторых действительно гомологичных элементов поведения животных и человека.
Весьма различной бывает и биологическая значимость проявлений «агрессивности» – от негативной до положительной, от патологии до адаптивной необходимости. Этим определяется важность широко осуществляемого в настоящее время изучения этих феноменов, в частности (при соблюдении должной осторожности при экстраполяции результатов исследования), для моделирования определенных психопатогенных состояний и ситуаций. Одно из центральных мест занимает сейчас эта тематика и при изучении этологических компонентов поведения человека, особенно в онтогенезе, в генетике поведения, а также в свете решений вопросов, связанных с проблемой социальных и биологических детерминант нашего поведения. В этих случаях «агрессивность» фигурирует главным образом как важный компонент фуппового (у животных) и социального (у человека) поведения.
Западноевропейские этологи внесли в свое время большой вклад в понимание значения поведения в процессе эволюции животного мира и в познании формирования видотипичного (инстинктивного) поведения в пределах зоологических систематических таксонов разного уровня.
Однако еще в 1941 г. Лоренц проанализировал кантовское априори для обоснования общебиологических концепций. Представления об аподиктичных научных истинах, предшествующих опыту и не зависящих от него, воплотились в практикуемый им бездоказательный, экспериментально ничем не обоснованный перенос этологических данных и выводов из мира животных в человеческое общество. Лоренц при этом пользовался лишь одними, притом чаще весьма сомнительными, аналогиями, сам же не осуществил ни одного эксперимента, в котором в сравнительном плане изучалось бы поведение животных и человека, а тем более доказывалась бы допустимость подобного рода экстраполяции.
Свое воплощение эта редукционистская тенденция находит и в глубоком теоретическом кризисе, переживаемом ныне зарубежной этологией, в частности так называемой «этологией человека». Первично изначальная предопределенность или предначертанность всего процесса эволюции поведения, «начало всех начал» преподносится при этом в разных вариантах. К.Лоренц, правда, уверяет, что не верит в существование сверхъестественных телеологических факторов, но еще меньше верит в действие одного-единственного «монокаузального» физиологического стимула. Он выдвигает постулат о некоей «телеономии», т.е. «сохраняющей вид целесообразности», которая хотя и модифицируется в результате селективного отбора, но в конечном итоге также определяется непознаваемыми первичными каузальностями. Аналогичные постулаты встречаются и у ряда последователей Лоренца, в частности у П.Лейхаузена.
Сформулированные Лоренцем взгляды составляют основной базис модной сейчас в Западной Европе «эволюционной теории познания». Ее приверженцы также являются преимущественно последователями и единомышленниками Лоренца. «Эволюционная теория познания» формировалась на основе «Альтенбергского семинара», организованного Лоренцем в своей резиденции под Веной. К ее «идейным отцам» относятся также английский неореалисг С.Александер, один из основоположников теории эмерджентной эволюции и известный австрийский философ К.Р.Поппер, а также Х.Альберт, в «критическом реализме» которого концепции Поппера получили дальнейшее развитие.
Как известно, в концепции эмерджентной эволюции сознание понимается как независимый от объекта компонент неореалистически понимаемого эволюционного процесса, как «поздний уровень» этого процесса. Достаточно напомнить, что эволюция, по Александеру, непредсказуема и научно необъяснима, а «низус» Александера – духовное начало, направляющее эволюцию к ее высшей цели – к божеству. Взгляды приверженцев эмерджентной эволюции основаны на концепциях неореализма, а поскольку последний тяготеет в гносеологии к субъективному идеализму, а в онтологии – к объективному идеализму и одновременно отрицает зависимость вещей от их взаимоотношений, то эта тенденция направлена на соединение науки и религии. Свидетельством тому является и метафизическая теория самодовлеющих «трех миров» Поппера (физического, ментального и объективного знания), как и его построенная на основе эмерджентизма теория сознания.
В полном соответствии с этими положениями «эволюционная теория познания» строится на представлениях о некоей «всеобщей» постепенной эмерджентной биологической эволюции, которая, по словам одного из ведущих идеологов этого течения профессора Венского университета Р. Ридля, представляет собой «континуум познавательных процессов, возраст которого равен возрасту жизни на этой планете». Инстинкты представляются при этом, как у Лоренца, извечно не изменяющимися, человеческая «способность к сознательному познанию», правда, постулируется как новейшее звено этого континуума, но даже человеческая речь представляется не специфически человеческим атрибутом, а лишь способностью к развитию общего с животными языка.
Проповедуя объективное познание «механизмов» биологической эволюции, сторонники этого направления объявляют ее причины вечно непознаваемыми. Приведя слова Канта из «Критики способности суждения», что конечная причина всегда была «чужаком в естествознании», и прибавляя, что таковой она осталась и по сегодняшний день, Ридль механически низводит цепь частных причин всеобщей эволюции к ее всеобщей первопричине, которая объявляется принципиально непознаваемой. При этом психика животных принимается за некое извечно замкнутое в себе, недетерминированное внутреннее начало, первопричинно предопределяющее все развития поведения и вообще всю жизнедеятельность животных. Таким образом, роль науки ограничивается здесь изучением лишь биологических феноменов поведения и их изменения в ходе филогенеза, поскольку познание каузальностей остается за пределами возможностей естественных наук.
Таким образом, «эволюционная теория познания» предстает перед нами как неприкрытая биологизация поведения человека: антропогенез, зарождение и развитие человеческого сознания, человеческого общества трактуются как всего-навсего незначительный эпизод и по своей сущности перманентно однозначной эволюции животного (!) мира. Логика этой концепции проста: не было и нет во всей истории живых существ иного процесса развития, кроме биологической эволюции, следовательно, природа человека и его поведение формировались на протяжении по меньшей мере тех 500 млн лет, которые потребовались для развития позвоночных животных.
Ясно одно: противопоставляется ли «инстинкт» «душе» или приравнивается к ней – результат один: при идеалистическом понимании первоисточников и движущих сил развития органического мира проблема происхождения и развития психики, факторов и направлений ее эволюции становится действительно неразрешимой. Вот к чему приводит новейшее извращение изучения эволюции психики, замена истинного научного поиска псевдонаучными метафизическими, идеалистическими спекуляциями.
* * *
Зоопсихологии приходится преодолевать и тенденцию упрощенчества, своего рода «физиологизацию», получившую сейчас особое распространение в связи с достижениями нейрофизиологии, биофизики, биохимии и других отраслей современной биологии. Эти тенденции сводятся к низведению психического к физиологическим и более элементарным процессам (например, рефлексам и таксисам) в сочетании с «выхолащиванием» содержания психической деятельности, с механистическим, биологизаторским игнорированием и просто отрицанием качественных отличий психики, психического отражения. К этому прибавляется еще непонимание (или нежелание понять) диалектики происхождения и развития психики в процессе эволюции, что составляет главным образом предмет зоопсихологии. Ошибка состоит в том, что в сфере психической деятельности животных целое сводится к частному, интегрированный комплекс – к составляющим его слагаемым («здание – к куче кирпичей»), высший порядок – к низшему (высший уровень биологической интеграции – к более низшему) и при всем этом стираются качественные различия между указанными уровнями и категориями.
Редукционистские тенденции проявляются подчас в стремлении (или попытках) «ликвидировать» зоопсихологию, как бы «отпеть» ее как научную дисциплину, провозглашая, что, дескать, ее предмет «растворился» в других науках, прежде всего – в нейрофизиологии. Такого рода посягательства предпринимались уже не раз и всегда были направлены не только против зоопсихологии, но неизбежно против всей психологии в целом. Выдающиеся психологи всегда понимали, что без зоопсихологии вообще невозможна психология как наука, во всяком случае если речь идет о марксистской психологии, строящей научный поиск на базе диалектико-материалистической методологии, исходя из понимания того, что психика человека не может быть научно достоверно познана вне изучения процесса ее развития.
Исторические корни редукционистских, биологизаторских тенденций в зоопсихологии, как и в общей психологии, простираются к началу нашего века, к знаменитым трудам Дж. Леба, автора теории тропизмов. Не ограничиваясь, однако, анализом элементарных форм поведения (преимущественно у низкоорганизованных животных), Леб пытался привести даже наиболее сложные поведенческие акты к элементарным реакциям – тропизмам (таксисам), а затем и к чисто физико-химическим явлениям. Механическими сентенциями и выпадами (разумеется, и против зоопсихологии) на уровне предельного редукционизма пестрят в наше время публикации Г. Темброка. На такой уровень опустился и К. Лоренц, который, также отрицая зоопсихологию, предпринял попытку, подобно Лебу (но с современным «обоснованием»), объяснить все формы поведения животных и человека элементарными таксисами и реакциями типа «обходного пути» (т.е. примитивнейшим решением задач в стимульной преградной ситуации). В этом же духе обсуждались, в частности, тенденции развития современных «наук о поведении» на недавно состоявшемся (в 1983 г.) в Берлине международном симпозиуме, посвященном «биологии поведения». С критикой биологи-заторских, редукционистско-нейрофизиологических тенденций в зоопсихологии и сравнительной психологии в последние годы выступили, в частности, Хольцкамп-Остеркамп, Барнетт, Бэрендс и другие исследователи.
* * *
Говоря о тенденциях современной зоопсихологии, следует упомянуть широко известные попытки установления языкового общения между человеком и шимпанзе. Это особая обширная тема, требующая специального обсуждения и не укладывающаяся в рамки этой публикации, но необходимо подчеркнуть, что главное здесь – отмежеваться от крикливого, сенсационного преподнесения результатов этих безусловно интересных опытов и дать им объективную научную оценку с позиций наших современных представлений о высших психических способностях животных, об их интеллекте. Напомним, что на эту цель были уже много лет тому назад направлены экспериментальные исследования советских зоопсихологов – Н.Н. Ладыгиной-Котс, впервые в истории ставившей психологические эксперименты на человекообразной обезьяне, Н.Ю. Войтониса, А.И. Каца, Н.А. Тих.
А.Г. Уланова вообще впервые (еще в 30-х годах) ввела в исследовательскую практику знаковое (подобно языку глухонемых) общение с обезьянами.
К сожалению, социализация поведения обезьян в биологизации поведения человека дискредитировали опыты Р. и Б. Гарднеров, Д. Премака, Д.М. Рамбо, Т. Гилла, П. Паттерсон и других по выявлению коммуникативных способностей антропоидов и возможностей «речевого» общения с ними. В свое время мы уже указывали на то, что, признавая безусловную ценность этих исследований, нельзя упускать из виду, что достигнутые результаты можно толковать не как свидетельство о естественных системах (и возможностях) антропоидов, а лишь как итоги дрессировки с применением подопытным животным сугубо человеческих, но не их собственных способов коммуникации. Принципиально эта искусственная система человеческих сигналов (символов, адекватных определенным обозначениям в языковом обиходе человека) ничем не отличается от постоянно практикуемого человеком в течение тысячелетий общения с разными одомашненными животными. Разумеется, общение человека с шимпанзе и количественно и качественно отличается от общения, скажем, с домашними животными, но суть дела от этого не меняется.
Множатся и выступления зарубежных ученых, ставящих под сомнение научную достоверность и эвристическую ценность обобщений, выведенных из опытов с «говорящими обезьянами», и указывающих на необходимость учета роли и значения глубинных структур и лингвистических универсалий в овладении подлинным языком, если иметь в виду человеческую речь.. Делается заключение, что поведение обезьян в этих опытах не имеет ничего общего с речью человека.
Способности обезьян к псевдоречевому общению с человеком, выработавшим у них этот артефакт путем дрессировки (путем «выбора на образец», введенного в практику зоопсихологических исследований еще более 60 лет тому назад Н.Н. Ладыгиной-Котс), стали для некоторых авторов, в том числе Д.Премака, мерилом интеллекта (или даже «сознания») этих животных. Некоторых исследователей упомянутые опыты стимулировали к выявлению у приматов способности к далеко идущему обобщению, формированию категорий и других форм интеллектуального поведения.
Вообще же фактические данные, полученные в опытах с «говорящими обезьянами», доказывают оправданность предположения, что у высших животных существуют генерализованные образы, из интеграции которых у каждого вида животных слагается некий, как, очевидно, можно его назвать, «генерализованный образ среды обитания». Этой формулировкой мы подчеркиваем сугубо биологическую сущность этой наивысшей, предельной для животных (и вместе с тем видеотипичной) формы психического отражения, ее избирательную ограниченность, фрагментарность, всецело обусловленную биологической валентностью компонентов занимаемой видом экологической ниши. Генерализованные образы животных функционально эквивалентны столь же гипотетическим пока «образам мира» у человека (по Леонтьеву), но, разумеется, качественно в корне отличаются от этой социально-исторически обусловленной категории.
Психические образы у животных – это процессы отражения, формирующиеся в обобщенном виде в результате накопления индивидуального опыта. Актуализация (хотя бы частичная) этого опыта происходит не только в уже знакомых, но и в новых, незнакомых ситуациях, в которых, однако, присутствуют элементы тех ситуаций, в которых эти образы формировались. В этом и заключается биологически адаптивное значение обобщенных психических образов. Проблема эта, конечно, не новая. В настоящее время ею (в общепсихологическим плане) занимаются особо С.Д. Смирнов и (в зоопсихологическом плане) Н.Н. Мешкова. Опираясь на свои экспериментальные исследования, посвященные ориентировочно-исследовательской деятельности животных, особенно в ситуациях новизны (т.е. при освоении нового пространственного окружения и при встрече с новыми предметами), Н.Н. Мешкова пришла к выводу, что формирующиеся у животных психические образы не только сохраняются, но непрерывно адаптивно изменяются и тем самым обеспечивают постоянную лабильную готовность животного к быстрым, даже неожиданным изменениям в окружающей среде.
Во многом очень близка советской зоопсихологии по тематике и методике подхода к решению задач психологического анализа поведения животных школа Б. Ренша. Представители этой школы сумели добыть весьма интересные, убедительные факты но различным вопросам, связанным с высшими психическими процессами у разных представителей позвоночных животных, в частности по авербальному мышлению и общению у животных, прямо пропорциональной зависимости между абсолютной величиной их мозга и психическими способностями, по манипулированию и гаптическим способностям животных как факторов психического развития и пр.
Особый интерес представляют работы по сравнительно-психологическому анализу движений передних конечностей и форм общения у приматов в свете антропогенеза. В этих работах усматривается тенденция к анализу морфофункциональных аспектов эволюции психики с учетом перехода количественных накоплений в качественные преобразования. Особенно созвучна советской школе зоопсихологов тенденция к выявлению значения манипулирования у животных как ведущего фактора эволюции психики.
* * *
Казалось бы, «академический» вопрос о психических образах у животных приобрел острое практическое значение, в этом, несомненно, состоит одна из важнейших, поистине новых тенденций зоопсихологии, можно даже сказать новая веха в истории этой науки.
Эта тенденция выросла постепенно из многолетнего участия психологов МГУ в различных отраслях практической деятельности человека: в народном хозяйстве (звероводство, животноводство), здравоохранении (дератизация, моделирование психопатогенных состояний и ситуаций), педагогике (дошкольное воспитание) и охране природы (программа «Экополис»). Достаточно напомнить, что раньше практическое значение зоопсихологии ощущалось разве что в служебном собаководстве и цирковой дрессировке.
Возникли объективные предпосылки и потребность разработки новой «общей прикладной зоопсихологии» с собственными задачами, предметом, концептуальным аппаратом, специфической методикой. Необходимость прикладных зоопсихологических исследований стала очевидной во всех случаях, когда обычно практикуемый подход на «чисто рефлекторном» уровне оказался неэффективным (это относится и к примитивно-этологическим методам работы). А такие ситуации возникают в практике все чаще, поскольку при таком подходе не охватывается поведение как единая, чрезвычайно сложная по своей структуре функция всего организма и, главное, не учитываются аспекты психического отражения, биопсихологические факторы его взаимоотношений со средой. Не менее важным участие зоопсихологов оказывается в тех случаях, когда в практике на передний план выступают сравнительно-психологические моменты, в частности общение человека с животными. Фундамент этого нового раздела заложен прикладными исследованиями зоопсихологов МГУ, отчасти уже внедренными в практику.
Особенно интенсивное развитие эта новая в зоопсихологии тенденция приобрела в связи с задачами, стоящими перед современным промышленным рыболовством. При этом тесная связь с практикой современного морского рыболовства привела к зарождению сразу двух разделов ихтиопсихологии: общей и прикладной.
Промышленное рыболовство переживает тяжелые времена: возможности экстенсивного его развития практически исчерпаны, интенсификация же его наталкивается на огромные трудности, одна из причин которых – слабая изученность поведения промысловых рыб в сочетании с игнорированием роли психической деятельности последних в прикладных ихтиологических исследованиях и конструкторских решениях, направленных на совершенствование процессов и средств лова. Необходимость учтения зоопсихологии в решении этой задачи сейчас уже признается специалистами промышленного рыболовства и выдвигается как настоятельное требование в официальных документах. Так, например, в решении Всесоюзной юбилейной научно-технической конференции по промышленному рыболовству, посвященной 100-летию со дня рождения основоположника науки о промышленном рыболовстве проф. Ф.И.Баранова, содержатся такие указания: «Чтобы научиться управлять поведением рыб, необходимо организовать его всестороннее изучение с помощью специалистов-зоопсихологов». Об объекте лова здесь говорится как о сложном организме с высоким уровнем психического отражения.
Ф.И.Баранов подверг всю проблематику морского рыболовства всестороннему анализу, вызывающему восхищение глубиной научного осмысления и эрудицией этого замечательного ученого. Итог этого титанического труда он сформулировал предельно кратко и четко: «... проблема трала упирается и перерастает в проблему поведения промысловых рыб».* Однако проводившиеся до сих пор исследования велись исключительно на элементарном рефлекторном или примитивно этологическом уровне и не дали ожидаемых результатов. Этот подход к изучению поведения животных (в данном случае рыб) потерпел фиаско при попытках внедрения в практику результатов лабораторных опытов.
* Баранов Ф.И. Техника промышленного рыболовства. М., 1960. С. 625.
Будучи приглашенными к участию в научных исследованиях по промышленному рыболовству, зоопсихологам МГУ пришлось поэтому прежде всего совместно со специалистами подвергнуть анализу бытующие ошибочные взгляды на процесс лова и противопоставить примитивному причинно-следственному пониманию поведения объектов лова такие основные положения разрабатываемого сейчас концептуального аппарата новой зоопсихологии, как, например, положения о ведущем адаптивном значении психического отражения в жизни животных (в частности, гидробионтов), об обобщенных психических образах как основе познавательных процессов у животных, о биологической и ситуационной обусловленности поведения животных и др.
Вместе с тем направление научного потока в сторону запросов промышленного рыболовства привело не только к необходимости создания новой отрасли зоопсихологии – уже упомянутой ихтиопсихологии, но и к началу работы по заполнению большого и очень значимого пробела к изучению эволюции психики: ведь рыбы являются родоначальниками всех позвоночных, а их психическая деятельность, психические компоненты их поведения практически, еще совершенно не исследованы. Более того, психическое отражение зародилось в воде и в течение половины срока существования животного мира (около 400 млн. из 800 (приблизительно) млн. лет) развивалось исключительно в водной среде. Уже этим определяется исключительная значимость изучения закономерностей психической деятельности зоологических гидробионтов.
Таким образом, начавшееся расширение поля деятельности зоопсихологов в сторону изучения вопросов, имеющих практическое значение, открывает также новые перспективы в изучении общетеоретических проблем, особенно по вопросам эволюции психики (а также ее онтогенетического развития). Этим в свою очередь создаются необходимые предпосылки для дальнейшей углубленной разработки и существенного обогащения всего концептуального аппарата новой зоопсихологии, а в итоге, в порядке обратной связи, и для уверенного подхода к решению многих новых крупномасштабных практических задач.
Учебное издание
фабри курт эрнестович
Основы зоопсихологии
Корректор Т.П.Толстова
Компьютерная верстка и дизайн: А.И.Чекалиной
Гарнитура: Times ET. Тираж 3000.464 с. Печать офсетная. Подписано в печать 07.12.1999. Издательство «Российское психологическое общество» ЛР №030651 от 14.07.1995
Содержание
TOC \o "1-3" Предисловие к 1-му изданию........................................................................................................................................................ PAGEREF _Toc519494969 \h 3
Предисловие ко 2-му изданию*.................................................................................................................................................. PAGEREF _Toc519494970 \h 3
К.Э.Фабри и зоопсихология в Московском государственном университете имени М. В. Ломоносова............................................................................................................................................................................................................................... PAGEREF _Toc519494971 \h 4
Введение......................................................................................................................................................................................................... PAGEREF _Toc519494972 \h 6
Предмет и задачи зоопсихологии...................................................................................................................................................... PAGEREF _Toc519494973 \h 6
Методы зоопсихологических исследований................................................................................................................................. PAGEREF _Toc519494974 \h 13
Часть I........................................................................................................................................................................................................... PAGEREF _Toc519494975 \h 18
ОБЩАЯ ХАРАКТЕРИСТИКА ПСИХИЧЕСКОЙ ДЕЯТЕЛЬНОСТИ ЖИВОТНЫХ.................................................................. PAGEREF _Toc519494976 \h 18
Глава 1 ПРОБЛЕМА ИНСТИНКТА И НАУЧЕНИЯ....................................................................................................................... PAGEREF _Toc519494977 \h 18
Ранние представления о психической деятельности животных........................................................................................... PAGEREF _Toc519494978 \h 18
Проблема инстинкта и научения в свете эволюционного учения........................................................................................... PAGEREF _Toc519494979 \h 21
Современное понимание проблемы инстинкта и научения....................................................................................................... PAGEREF _Toc519494980 \h 26
Глава 2 ИНСТИНКТИВНОЕ ПОВЕДЕНИЕ........................................................................................................................................ PAGEREF _Toc519494981 \h 29
Инстинктивное поведение как основа жизнедеятельности животных.............................................................................. PAGEREF _Toc519494982 \h 29
Внутренние факторы инстинктивного поведения..................................................................................................................... PAGEREF _Toc519494983 \h 30
Внешние факторы инстинктивного поведения........................................................................................................................... PAGEREF _Toc519494984 \h 33
Структура инстинктивного поведения......................................................................................................................................... PAGEREF _Toc519494985 \h 37
Инстинктивное поведение и общение............................................................................................................................................. PAGEREF _Toc519494986 \h 40
Психический компонент инстинктивного поведения................................................................................................................. PAGEREF _Toc519494987 \h 44
Глава 3 НАУЧЕНИЕ............................................................................................................................................................................... PAGEREF _Toc519494988 \h 45
Общая характеристика процесса научения................................................................................................................................. PAGEREF _Toc519494989 \h 45
Навык....................................................................................................................................................................................................... PAGEREF _Toc519494990 \h 48
Научение и общение. Подражание.................................................................................................................................................. PAGEREF _Toc519494991 \h 56
Часть II.......................................................................................................................................................................................................... PAGEREF _Toc519494992 \h 59
РАЗВИТИЕ ПСИХИЧЕСКОЙ ДЕЯТЕЛЬНОСТИ ЖИВОТНЫХ В ОНТОГЕНЕЗЕ..................................................................... PAGEREF _Toc519494993 \h 59
Глава 1 ПРОБЛЕМА ОНТОГЕНЕЗА ПОВЕДЕНИЯ........................................................................................................................ PAGEREF _Toc519494994 \h 59
Врожденное и приобретаемое в индивидуальном развитии поведения.............................................................................. PAGEREF _Toc519494995 \h 59
Биологическая обусловленность онтогенеза поведения животных..................................................................................... PAGEREF _Toc519494996 \h 60
Глава 2 РАЗВИТИЕ ПСИХИЧЕСКОЙ ДЕЯТЕЛЬНОСТИ В ПРЕНАТАЛЬНОМ ПЕРИОДЕ.................................................. PAGEREF _Toc519494997 \h 61
Врожденное и приобретаемое в пренатальном развитии поведения.................................................................................. PAGEREF _Toc519494998 \h 62
Сравнительный обзор развития двигательной активности зародышей............................................................................ PAGEREF _Toc519494999 \h 65
Пренатальное развитие сенсорных способностей и элементов общения........................................................................... PAGEREF _Toc519495000 \h 70
Эмбриогенез и развитие психического отражения.................................................................................................................. PAGEREF _Toc519495001 \h 72
Глава 3 РАЗВИТИЕ ПСИХИЧЕСКОЙ ДЕЯТЕЛЬНОСТИ В РАННЕМ ПОСТНАТАЛЬНОМ ПЕРИОДЕ............................ PAGEREF _Toc519495002 \h 73
Особенности постнатального развития поведения животных............................................................................................ PAGEREF _Toc519495003 \h 73
Инстинктивное поведение в раннем постнатальном периоде................................................................................................ PAGEREF _Toc519495004 \h 77
Ранний опыт.......................................................................................................................................................................................... PAGEREF _Toc519495005 \h 81
Взаимоотношения между компонентами раннего постнатального развития поведения............................................. PAGEREF _Toc519495006 \h 88
Раннее формирование общения........................................................................................................................................................ PAGEREF _Toc519495007 \h 91
Познавательные аспекты раннего постнатального поведения............................................................................................. PAGEREF _Toc519495008 \h 93
Глава 4 РАЗВИТИЕ ПСИХИЧЕСКОЙ ДЕЯТЕЛЬНОСТИ В ЮВЕНИЛЬНОМ (ИГРОВОМ) ПЕРИОДЕ............................... PAGEREF _Toc519495009 \h 98
Общая характеристика игры у животных.................................................................................................................................. PAGEREF _Toc519495010 \h 98
Совершенствование двигательной активности в играх животных................................................................................. PAGEREF _Toc519495011 \h 101
Формирование общения в играх животных............................................................................................................................... PAGEREF _Toc519495012 \h 104
Познавательная функция игровой активности животных................................................................................................... PAGEREF _Toc519495013 \h 107
Часть III...................................................................................................................................................................................................... PAGEREF _Toc519495014 \h 111
ЭВОЛЮЦИЯ ПСИХИКИ.......................................................................................................................................................................... PAGEREF _Toc519495015 \h 111
Вводные замечания............................................................................................................................................................................. PAGEREF _Toc519495016 \h 111
Глава 1 ЭЛЕМЕНТАРНАЯ СЕНСОРНАЯ ПСИХИКА................................................................................................................. PAGEREF _Toc519495017 \h 114
Низший уровень психического развития..................................................................................................................................... PAGEREF _Toc519495018 \h 114
Высший уровень развития элементарной сенсорной психики.............................................................................................. PAGEREF _Toc519495019 \h 123
Глава 2 ПЕРЦЕПТИВНАЯ ПСИХИКА............................................................................................................................................. PAGEREF _Toc519495020 \h 133
Низший уровень развития перцептивной психики................................................................................................................... PAGEREF _Toc519495021 \h 134
Высший уровень развития перцептивной психики.................................................................................................................. PAGEREF _Toc519495022 \h 148
Проблема интеллекта животных................................................................................................................................................. PAGEREF _Toc519495023 \h 159
Глава 3 ЭВОЛЮЦИЯ ПСИХИКИ И АНТРОПОГЕНЕЗ................................................................................................................ PAGEREF _Toc519495024 \h 167
Проблема происхождения трудовой деятельности............................................................................................................... PAGEREF _Toc519495025 \h 168
Проблема зарождения общественных отношений и членораздельной речи.................................................................... PAGEREF _Toc519495026 \h 176
Литература......................................................................................................................................................................................... PAGEREF _Toc519495027 \h 182
Приложение.......................................................................................................................................................................................... PAGEREF _Toc519495028 \h 183
Игры животных и игры детей (сравнительно-психологические аспекты)*.................................................................. PAGEREF _Toc519495029 \h 183
Научное наследие А.Н. Леонтьева и вопросы эволюции психики*....................................................................................... PAGEREF _Toc519495030 \h 191
Зоопсихология*................................................................................................................................................................................... PAGEREF _Toc519495031 \h 202
Содержание............................................................................................................................................................................................. PAGEREF _Toc519495032 \h 210